Хорион

1 - маточная труба, 2 - decidua basalis, 3 - decidua capsularis, 4 - миометрий, 5 - влагалище, 6 - ворсинки хорионального мешка, 7 - decidua parietalis, 8 - полость хориона, 9 - полость матки, 10 - амнион, 11 - амниотическая полость, 12 - пространство между ворсинками, 13 - ворсинки хориона, 14 - гладкий хорион, 15 - внутренний зев
Амнион Амнион образует заполненный жидкостью мембранозный амниотический мешок, который окружает эмбрион (Рис.2 Б). Амнион соединен с краями эмбрионального диска. Стрелками показаны направления роста амниотической полости. Место соединения амниона с эмбриональным диском в процессе формирования головной, каудальной и латеральных складок смещается на вентральную поверхность эмбриона (Рис.1Б). Первоначально амниотическая жидкость секретируется амниотическими клетками, но постепенно все большая ее часть возникает из материнской крови, транспортируемой через амнион от париетальной части децидуальной оболочки (decidua parietalis) и межворсинчатых пространств плаценты.

Рис. 2. Процесс покрытия амнионом пупочного канатика и частичного вовлечения желточного мешка в первичную кишку эмбриона. Показано также образование плодной части плаценты и дегенерация ворсинок хориона. А - 3 недели; Б - 4 недели; В - 10 недель; Г - 20 недель
1 - амнион, 2 - соединяющий стебелек, 3 - эмбриональный диск, 4 - желточный мешок, 5 - хорион, 6 - стебелек хориона, 7 - полость хориона, 8 - амниотический мешок, 9 - пупочный канатик, 10 - остатки желточного мешка, 11 - гладкий хорион
Эмбрион 
Рис. 3. А и Б - зародыш человека длиной 4 мм. Голова далеко отстоит от хвоста и прилегает к сердечному выступу. Зачатки глаз в виде небольших круглых пятнышек. Хорошо выражены жаберные дуги и щели. Зачатки верхних конечностей на уровне 8-12 сомитов. Эмбрион примерно 28 дней развития. В - сагитальный разрез того же эмбриона
1 - , 2 - , 3 - мозговые пузыри - передний, средний и задний, 4 - стомодеум, 5 - карман Ратке, 6 - зачаток языка, 7 - сердце, 8 - печень, 9 - брюшная аорта, 10 - зачаток спинного мозга, 11 - пищевод, 12 - трехея
Fig. 3. А и Б - Embryo, crown-rump lengh - 4 mm. The head is far distant from the tail and adjoins to the heart prominence. The optical primordia have the form of smoll round spots seen through the skin. Branchial arches and grooves are well pronounced. There are primordia of upper limbs in the form of smoll eminences located at the level of 8-12 somites. By its appearance, the number of somites and the size of embryo resembles the 30-days embryo. B - sagital section of the embryo
1 - , 2 - , 3 - brain vesicle - anterior, middle and posterior , 4 - stomodeum, 5 - Rathke's pouch, 6 - tongue primordium, 7 - heart, 8 - liver, 9 - abdominal aorta, 10 - primordium of the spinal cord, 11 - oesophagus, 12 - trachea
Эмбрион человека, 28-й день развития. Эмбрион имеет С-образный изгиб. Каудальный нейропор закрывается. Зачатки верхних конечностей становятся похожими на плавники. Видны 4 пары бранхиальных дуг. Появляются зачатки нижних конечностей. Формируются слуховые пузырьки. Видны хрусталиковые плакоды, эктодермальные утолщения для будущих хрусталико глаз.
Хвост уменьшается, но все еще является характерным признаком эмбриона
Формирование лица
На каждой стороне лобно-носового выпячивания возникает овальное утолщение поверхностной эктодермы, называемое носовой плакодой (nasal placodes) (Рис.2, 3)
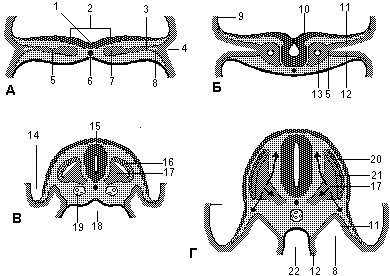
Образование эмбриональных складок Заканчивается образование головной складки. Зачаток сердца и перикардиальная полость оказываются внутри эмбриона (Рис 3.Б). Хвостовая скадка, возникшая вначале в результате роста спинного мозга в каудальном направлении углубляется в кранио-дорсальном направлении в результате каудальная часть эмбриона оказывается над клоачной мембраной (Рис.1) В это же время часть желточного мешка включается в эмбрион как задняя кишка (hindgut) (зачаток нисходящего колона). Терминальная часть задней кишки слегка расширяется и формирует клоаку ( зачаток мочевого пузыря и прямой кишки). После окончания образования хвостовой складки остаток примитивной полоски оказывается теперь расположенным не краниальнее, а каудальнее клоачной мембраны. 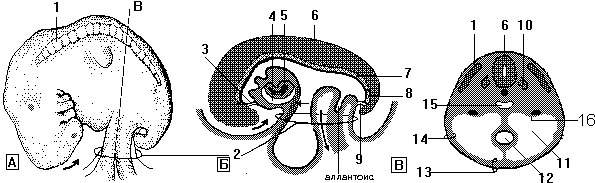
Рис. 5. Эмбрион человека, 28-й день развития. А - внешний вид эмбриона сбоку, амнион удален. Б - сагитальный разрез, показаны составляющие пупочного канатика. В - поперечный разрез по штриховой линии, показанной на схеме Б.
1 - сомит, 2 - пупочный канатик, 3 - орофарингеальная мембрана, 4 - перикардиальная полость, 5 - сердце, 6 - нервная трубка, 7 - хорда, 8 - остатки примитивной полоски, 9 - клоачная мембрана, 10 - ганглии задних корешков спинальных нервов, 11 - внутриэмбриональный целом, 12 - средняя кишка, 13 - вентральная стенка живота, 14 - латеральная стенка живота, 15 - аорта, 16 - урогенитальный гребень Латеральные складки смыкаются по медиальной плоскости, за исключением уровня желточного стебля, устье которого сильно сужается. Область присоединения амниона к эмбриону в целом также сильно сужается , превращаясь в пупочную область (umbilical cord) (Рис.4.Б). Смыкаясь латеральные складки уменьшают область сообщения внутриэмбрионального целома с внеэмбриональным целомом до узкого канала, который сохраняется вплоть до конца 10-й недели (длинная стрелка на рис.2Б).
Примитивная полоска
Примитивная полоска заканчивает активное формировние внутриэмбриональной мезодермы.
Хорда
Хорда представлена непрерывным цилиндром клеток, идущим через развивающийся позвоночный столб. Плотно упакованные клетки хорды содержат значительные отложения гликогена и цитофиламент. Клетки связаны многочисленными десмосомами . Аморфная мембрана вокруг хорды утолщается начинает походить на оболочку,состоящую из тоноко филаментозного материала. Плотность клеток в аксиальной мезенхиме увеличивается.
Брыжейки В результате образования латеральных эмбриональных складок и движения их навстречу друг другу ножки или латеральные части внутриэмбрионального целома также смещаются навстеречу друг другу и в вентральной части эмбриона они встречаются, разделенные дорзальнее и вентральнее сердца и кишки только двумя слоями спланхнической мезодермы (мезокардием и мезентерием) (Рис.4 В). В задней части эмбриона вентральный мезентерий начинает дегенерировать, в результате здесь образуется большая эмбриональная перитонеальная полость. Становятся хорошо различимы три целомические полости: (1) большая перикардиальная полость (результат дегенерации вентрального мезокардия), (2) два относительно небольших перикардиоперитонеальных канала (pericardioperitoneal canals) и (3) большая перитонеальная полость ( результат дегенерации вентрального мезентерия).
Мезентерий желудка.
На данной стадии развития желудок расположен в срединной плоскости и связан с дорзальной стенкой брюшной полости дорзальным мезентерием (dorsal mesentery) часто называемым дорзальным мезогастрием (dorsal mesogastrium), а с вентральной брюшной стенкой он связан вентральным мезентерием или вентральным мезогастрием (Рис. 4 )
Экспансия амниотической полости ведет к облитерации внеэмбриональнго целома и амнинон начинает формировать эпителиальную оболочку для пупочного канатика.
 Продолжается процесс сомитогенеза.
Продолжается процесс сомитогенеза.
Рис. 6 г. Поперечные срезы через эмбрион человека. А - пресомитный эмбрион (около 18 дней),видна параксиальная мезодерма, из которой происходят сомиты. Б - эмбрион около 22 дней. В - эмбрион около 26 дней. Область дермомиотома сомита дает миотом и дерматом (будущая дерма), Г - эмбрион около 28 дней. Стрелками показана миграция клеток из области склеротома сомита
1 - нервная борозда, 2 - нервная пластинка, 3 - латеральная мезодерма, 4 - внеэмбриональный целом, 5 - промежуточная мезодерма, 6 - хорда, 7 - параксиальная мезодерма, 8 - внутриэмбриональный целом, 9 - амнион, 10 - нервная складка, 11 - соматическая мезодерма, 12 - спланхническая мезодерма, 13 - сомит, 14 - амниотическая полость, 15 - нервная трубка, 16 - дермомиотом, 17 - склеротом, 18 - желточный мешок, 19 - дорсальная аорта, 20 - дерматом, 21 - миотом, 22 - первичная кишка
Клетки нервного гребеняКлетки нервного гребня дают специфические скелетные структуры, например, эти клетки первой жаберной дуги дают Меккелев хрящ, верхнюю и нижнюю челюсть, различные связки и 2 из 3-х малых косточек, оссиклей, среднего уха. Типичная бранхиальная дуга содержит артерию (aortic arch), полоску хряща, мышечный компонент и нерв. (Рис.6.Б, Г-Е).
Жаберный аппарат Развитие жаберных дуг начинается с миграции клеток нервного гребня (neural crest cells) в область будущей головы и шеи. В результате проникновения этих клеток образуется незначительное возвышение по бокам развивающейся глотки, первая бранхиальная дуга, зачаток челюстей. На 24-й день развития обнаруживается появление второй жаберной дуги , на 26-й день - третьей и на 28-й день - четвертой жаберной дуги. Ч-я и 6-я жаберные дуги малы и не могут быть видны на поверхности эмбриона. Жаберные дуги отделяются друг от друга хорошо заметными щелями, называемыми жаберными щелями (branchial grooves) (Рис.5А)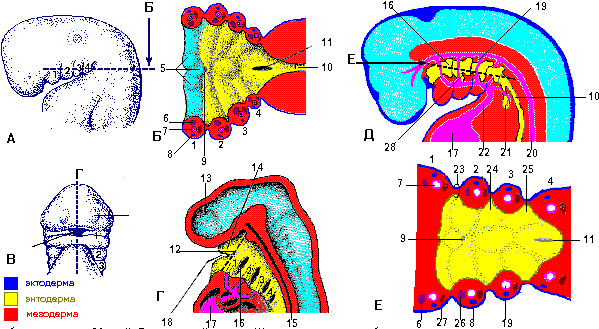
Рис. 7. Эмбрион человека (28 дней). Бранхиальный аппарат. Штриховыми линиями обозначены уровни разрезов
1 - , 2 - , 3 - , 4 - 1-4-я бранхиальные дуги, 5 - зачаток языка, 6 - Меккелев хрящ, 7 - первая дуга аорты, 8 - нерв, 9 - слепое отверстие, 10 - пищевод, 11 - ларинготрахеальная борозда, 12 - положение орофарингеальной мембраны, 13 - передний мозг, 14 - верхнечелюстное выпячивание, 15 - хорда, 16 - 1-й глоточный карман, 17 - сердце, 18 - рот, 19 - 3-я дуга аорты, 20 - дорсальная аорта, 21 - зачаток легкого, 22 - truncus arteriosus, 23 - 1-я бранхиальная мембрана, 24, 25 - 2-й и 3-й глоточные карманы, 26 - хрящ Рейхарта, 27 - мышцы, 28 - тироидный дивертикул
Первая жаберная дуга часто называется нижнечелюстной дугой (mandibular arch), она состоит из двух выпячиваний ("отростков") : (1) большого нижнечелюстного выпячивания (mandibular prominence), формирующего нижнюю челюсть, и (2) небольшого верхнечелюстного выпячивания ( maxillary prominence), формирующего верхнюю челюсть, скуловую кость (zygomatic bone) и чешуйчатую часть височной кости (tempotal bone) (Рис.7.В).
Вторая жаберная дуга часто называется гиоидной (hyoid arch), так как вносит вклад в образование подъязычной кости. Остальные жаберные дуги обозначаются только номерами.
Мезенхима каждой бранхиальной дуги дает начало мышцам, хрящам и кости.
Клетки нервного гребня дают специфические скелетные структуры, например, эти клетки первой жаберной дуги дают Меккелев хрящ, верхнюю и нижнюю челюсть, различные связки и 2 из 3-х малых косточек, оссиклей, среднего уха. Типичная бранхиальная дуга содержит артерию (aortic arch), полоску хряща, мышечный компонент и нерв. (Рис.5.Б, Г-Е).
Кровь, проходящая через аортальные дуги вступает в дорзальную аорту (Рис.6.Д,Г). 
Рис. 8. Рис. 8. А - область головы и шеи эмбриона человека (28 дней). Показаны зачатки мышц бранхиальных дуг. Стрелка показыват путь миграции миобластов из окципитальных миотомов для образования мускулатуры языка. Б - голова и шея плода человека ( 20 недель). Показаны мышцы, происходящие из бранхиальных дуг ( часть мышц platisma и sternocleidomastoid удалена, чтобы показать более глубокие мышцы). Миобласты из 2-й бранхиальной дуги мигрируют в область шеи и дают мышцы лица
1 - затылочные миотомы, 2 - m. orbicularis, 3 - m. buccinator, 4 - m. orbitalis oculi, 5 - m. frontalis, 6 - m. temporalis, 7 - v. occipitalis, 8 - m. auricularis, 9 - m. stylohyoid, 10 - m. stylopharingeus, 11 - anterior and posterior bellies of digastric, 12 - m. platisma, 13 - ключица, 14 - m. sternocleidomastoid, 15 - pharingeal muscles, 16 - m. mylohyoid, 17 - m. masseter
Хрящи бранхиальных дуг. Хрящевой компонент 1-й жаберной дуги называется Меккелевым хрящем (Meckel's cartilage), 2-й дуги - хрящем Рейхерта (Reichert's cartilage). Хрящи 3-й и 4-й дуг не имеют специального названия (см.29-й день, Рис.6). На рис.2 показано положение мышечных компонентов жаберных дуг (Рис.7.А) и их производных на голове и шее 20-недельного плода (Рис.7.Б).
А - вид сбоку на область головы и шеи у эмбриона человека 4-х недель. Показаны черепно-лицевые нервы, производные соответствующих бранхиальных дуг. Б - голова плода (20 недель). Показано поверхностное распределение двух каудальных ветвей тройничного нерва (V), производных первой бранхиальной дуги. В - сагитальный разрез через голову плода. Показано распределение глубоких сенсорных волокон бранхъиальных нервов к зубам, слизистой языка, глотке, полости носа, нёбу и гортани
1 - язык, 2 - нёбо, 3 - полость носа, 4 - паратидная железа, 5 - глотка, 6 - пищевод, 7 - гортань, 8 - низнечелюстная железа, 9 - подъязычные железы.
Состав внеклеточного матрикса
Компонент Хорда Аксиальная
мезенхима
Kлетки Оболочка 0 - отсутствие, (+) - слабая, + - сильная, ++ - очень сильная реакция
* - экспрессия, по-видимому, ограничена базальной мембраной Ходроитин/
дерматан сульфат 0 ++ +
Кератан сульфат 0 0 0
Гиалуроновая кислота 0 + +
Аггрекан 0 0 0
Бигликан + 0 0
Коллаген тип I (+), + (+) + <
Коллаген тип II 0 + 0
Декорин 0 ++ +
Фибронектин 0 ++ (+)
Ламинин (+) +* (+)
Тенасцин 0 + 0
TGF-β + > 0 0
Иммунореактивность и интенсивность окрашивания на 'TGF-beta увеличивается
Протеогликан Декорин обычно взаимодействует с коллагеновыми фибриллами, т,е.
участвует в фибриллогенезе, роль бигликана неизвестна. Коллаген тип II,по-видимому,
участвует в эпителиально-мезенхимных взаимодействиях (оболочка). Гликопротеин
фибронектин, по-видимому, связан с базальной мембраной и может выполнять
индуктивную роль. Молекулы ламинина, по-видимому, локализуются в
эндоплазматическом ретикулеме или аппарате Гольджи. Появление тенасцина в
оболочке хорды, по-видимому, связано с хондрогенезом зачатков тел позвонков и
межпозвоночных дисков, испытывающих преходящую хрящевую дифференцировку.
Секреция почти всех компонентов ВКМ в хорде человека регулируется TGF-beta. Так
TGF-beta индуцирует синтез декорина, который может связываться с фактором роста
через его core белок, модулируя его активность. Сходный механизм негативной петли
обратной связи обнаружен для ламинина.
В аксиальной мезенхиме межклеточные пространства заполнены
хондроитин/дерматан сульфатом и гиалуроновой кислотой. Они, особенно последняя,
ответственны за формирование гидратированных пространств, способбствующих
экспансии ВКМ ,миграции клеток и ингибированию дифференцировки или
хондрогенеза. След., склеротомные клетки иммигрирующие в аксиальную область
находят здесь идеальные условия для миграции и нахождения финальной позиции.
Этому способствует и фибронектин ВКМ аксиальной мезенхимы(Gotz et al., 1995).
Эмбрионы кур (Ст.) Эмбрионы мыши (Ст.)По Theiler's (1989) 16 стадия развития ,10-й день
В кровяных островках,образующихся на ранней сомитной стадии (Е8.0) ангиобласты все еще обнаруживают высокий уровень экспрессии flt-1, тогда как большинство гематопоэтических клеток экспрессирует flt-1
на низком уровне. Далее экспрессия flt-1 в гематопоэтических клетках снижается и наконец, экспрессия ограничивается только эндотелиальными клетками , достигая максимума к среднесомитной стадии (Е8.5). Это указывает на то, что гематопоэтические и эндотелиальные клетки происходят из общего предшественника. Затем экспрессия flt-1 в эндотелиальных клетках снижается и поддерживается на очень низком уровне на поздних стадиях эмбриогенеза.
1 - амнион, 2 - соединяющий стебелек, 3 - эмбриональный диск, 4 - желточный мешок, 5 - хорион, 6 - стебелек хориона, 7 - полость хориона, 8 - амниотический мешок, 9 - пупочный канатик, 10 - остатки желточного мешка, 11 - гладкий хорион
Эмбрион Рис. 3. А и Б - зародыш человека длиной 4 мм. Голова далеко отстоит от хвоста и прилегает к сердечному выступу. Зачатки глаз в виде небольших круглых пятнышек. Хорошо выражены жаберные дуги и щели. Зачатки верхних конечностей на уровне 8-12 сомитов. Эмбрион примерно 28 дней развития. В - сагитальный разрез того же эмбриона
1 - , 2 - , 3 - мозговые пузыри - передний, средний и задний, 4 - стомодеум, 5 - карман Ратке, 6 - зачаток языка, 7 - сердце, 8 - печень, 9 - брюшная аорта, 10 - зачаток спинного мозга, 11 - пищевод, 12 - трехея
Fig. 3. А и Б - Embryo, crown-rump lengh - 4 mm. The head is far distant from the tail and adjoins to the heart prominence. The optical primordia have the form of smoll round spots seen through the skin. Branchial arches and grooves are well pronounced. There are primordia of upper limbs in the form of smoll eminences located at the level of 8-12 somites. By its appearance, the number of somites and the size of embryo resembles the 30-days embryo. B - sagital section of the embryo
1 - , 2 - , 3 - brain vesicle - anterior, middle and posterior , 4 - stomodeum, 5 - Rathke's pouch, 6 - tongue primordium, 7 - heart, 8 - liver, 9 - abdominal aorta, 10 - primordium of the spinal cord, 11 - oesophagus, 12 - trachea
1 - , 2 - , 3 - мозговые пузыри - передний, средний и задний, 4 - стомодеум, 5 - карман Ратке, 6 - зачаток языка, 7 - сердце, 8 - печень, 9 - брюшная аорта, 10 - зачаток спинного мозга, 11 - пищевод, 12 - трехея
Fig. 3. А и Б - Embryo, crown-rump lengh - 4 mm. The head is far distant from the tail and adjoins to the heart prominence. The optical primordia have the form of smoll round spots seen through the skin. Branchial arches and grooves are well pronounced. There are primordia of upper limbs in the form of smoll eminences located at the level of 8-12 somites. By its appearance, the number of somites and the size of embryo resembles the 30-days embryo. B - sagital section of the embryo
1 - , 2 - , 3 - brain vesicle - anterior, middle and posterior , 4 - stomodeum, 5 - Rathke's pouch, 6 - tongue primordium, 7 - heart, 8 - liver, 9 - abdominal aorta, 10 - primordium of the spinal cord, 11 - oesophagus, 12 - trachea

Эмбрион имеет С-образный изгиб. Каудальный нейропор закрывается. Зачатки верхних конечностей становятся похожими на плавники. Видны 4 пары бранхиальных дуг. Появляются зачатки нижних конечностей. Формируются слуховые пузырьки. Видны хрусталиковые плакоды, эктодермальные утолщения для будущих хрусталико глаз.
Хвост уменьшается, но все еще является характерным признаком эмбриона
Формирование лица
На каждой стороне лобно-носового выпячивания возникает овальное утолщение поверхностной эктодермы, называемое носовой плакодой (nasal placodes) (Рис.2, 3)
Образование эмбриональных складок Заканчивается образование головной складки. Зачаток сердца и перикардиальная полость оказываются внутри эмбриона (Рис 3.Б). Хвостовая скадка, возникшая вначале в результате роста спинного мозга в каудальном направлении углубляется в кранио-дорсальном направлении в результате каудальная часть эмбриона оказывается над клоачной мембраной (Рис.1) В это же время часть желточного мешка включается в эмбрион как задняя кишка (hindgut) (зачаток нисходящего колона). Терминальная часть задней кишки слегка расширяется и формирует клоаку ( зачаток мочевого пузыря и прямой кишки). После окончания образования хвостовой складки остаток примитивной полоски оказывается теперь расположенным не краниальнее, а каудальнее клоачной мембраны.
Рис. 5. Эмбрион человека, 28-й день развития. А - внешний вид эмбриона сбоку, амнион удален. Б - сагитальный разрез, показаны составляющие пупочного канатика. В - поперечный разрез по штриховой линии, показанной на схеме Б.
1 - сомит, 2 - пупочный канатик, 3 - орофарингеальная мембрана, 4 - перикардиальная полость, 5 - сердце, 6 - нервная трубка, 7 - хорда, 8 - остатки примитивной полоски, 9 - клоачная мембрана, 10 - ганглии задних корешков спинальных нервов, 11 - внутриэмбриональный целом, 12 - средняя кишка, 13 - вентральная стенка живота, 14 - латеральная стенка живота, 15 - аорта, 16 - урогенитальный гребень Латеральные складки смыкаются по медиальной плоскости, за исключением уровня желточного стебля, устье которого сильно сужается. Область присоединения амниона к эмбриону в целом также сильно сужается , превращаясь в пупочную область (umbilical cord) (Рис.4.Б). Смыкаясь латеральные складки уменьшают область сообщения внутриэмбрионального целома с внеэмбриональным целомом до узкого канала, который сохраняется вплоть до конца 10-й недели (длинная стрелка на рис.2Б).
Примитивная полоска
Примитивная полоска заканчивает активное формировние внутриэмбриональной мезодермы.
Хорда
Хорда представлена непрерывным цилиндром клеток, идущим через развивающийся позвоночный столб. Плотно упакованные клетки хорды содержат значительные отложения гликогена и цитофиламент. Клетки связаны многочисленными десмосомами . Аморфная мембрана вокруг хорды утолщается начинает походить на оболочку,состоящую из тоноко филаментозного материала. Плотность клеток в аксиальной мезенхиме увеличивается.
Брыжейки В результате образования латеральных эмбриональных складок и движения их навстречу друг другу ножки или латеральные части внутриэмбрионального целома также смещаются навстеречу друг другу и в вентральной части эмбриона они встречаются, разделенные дорзальнее и вентральнее сердца и кишки только двумя слоями спланхнической мезодермы (мезокардием и мезентерием) (Рис.4 В). В задней части эмбриона вентральный мезентерий начинает дегенерировать, в результате здесь образуется большая эмбриональная перитонеальная полость. Становятся хорошо различимы три целомические полости: (1) большая перикардиальная полость (результат дегенерации вентрального мезокардия), (2) два относительно небольших перикардиоперитонеальных канала (pericardioperitoneal canals) и (3) большая перитонеальная полость ( результат дегенерации вентрального мезентерия).
Мезентерий желудка.
На данной стадии развития желудок расположен в срединной плоскости и связан с дорзальной стенкой брюшной полости дорзальным мезентерием (dorsal mesentery) часто называемым дорзальным мезогастрием (dorsal mesogastrium), а с вентральной брюшной стенкой он связан вентральным мезентерием или вентральным мезогастрием (Рис. 4 )
Экспансия амниотической полости ведет к облитерации внеэмбриональнго целома и амнинон начинает формировать эпителиальную оболочку для пупочного канатика.
Продолжается процесс сомитогенеза.
Рис. 6 г. Поперечные срезы через эмбрион человека. А - пресомитный эмбрион (около 18 дней),видна параксиальная мезодерма, из которой происходят сомиты. Б - эмбрион около 22 дней. В - эмбрион около 26 дней. Область дермомиотома сомита дает миотом и дерматом (будущая дерма), Г - эмбрион около 28 дней. Стрелками показана миграция клеток из области склеротома сомита
1 - нервная борозда, 2 - нервная пластинка, 3 - латеральная мезодерма, 4 - внеэмбриональный целом, 5 - промежуточная мезодерма, 6 - хорда, 7 - параксиальная мезодерма, 8 - внутриэмбриональный целом, 9 - амнион, 10 - нервная складка, 11 - соматическая мезодерма, 12 - спланхническая мезодерма, 13 - сомит, 14 - амниотическая полость, 15 - нервная трубка, 16 - дермомиотом, 17 - склеротом, 18 - желточный мешок, 19 - дорсальная аорта, 20 - дерматом, 21 - миотом, 22 - первичная кишка
Клетки нервного гребеняКлетки нервного гребня дают специфические скелетные структуры, например, эти клетки первой жаберной дуги дают Меккелев хрящ, верхнюю и нижнюю челюсть, различные связки и 2 из 3-х малых косточек, оссиклей, среднего уха. Типичная бранхиальная дуга содержит артерию (aortic arch), полоску хряща, мышечный компонент и нерв. (Рис.6.Б, Г-Е).
Жаберный аппарат Развитие жаберных дуг начинается с миграции клеток нервного гребня (neural crest cells) в область будущей головы и шеи. В результате проникновения этих клеток образуется незначительное возвышение по бокам развивающейся глотки, первая бранхиальная дуга, зачаток челюстей. На 24-й день развития обнаруживается появление второй жаберной дуги , на 26-й день - третьей и на 28-й день - четвертой жаберной дуги. Ч-я и 6-я жаберные дуги малы и не могут быть видны на поверхности эмбриона. Жаберные дуги отделяются друг от друга хорошо заметными щелями, называемыми жаберными щелями (branchial grooves) (Рис.5А)
Рис. 7. Эмбрион человека (28 дней). Бранхиальный аппарат. Штриховыми линиями обозначены уровни разрезов
1 - , 2 - , 3 - , 4 - 1-4-я бранхиальные дуги, 5 - зачаток языка, 6 - Меккелев хрящ, 7 - первая дуга аорты, 8 - нерв, 9 - слепое отверстие, 10 - пищевод, 11 - ларинготрахеальная борозда, 12 - положение орофарингеальной мембраны, 13 - передний мозг, 14 - верхнечелюстное выпячивание, 15 - хорда, 16 - 1-й глоточный карман, 17 - сердце, 18 - рот, 19 - 3-я дуга аорты, 20 - дорсальная аорта, 21 - зачаток легкого, 22 - truncus arteriosus, 23 - 1-я бранхиальная мембрана, 24, 25 - 2-й и 3-й глоточные карманы, 26 - хрящ Рейхарта, 27 - мышцы, 28 - тироидный дивертикул
Первая жаберная дуга часто называется нижнечелюстной дугой (mandibular arch), она состоит из двух выпячиваний ("отростков") : (1) большого нижнечелюстного выпячивания (mandibular prominence), формирующего нижнюю челюсть, и (2) небольшого верхнечелюстного выпячивания ( maxillary prominence), формирующего верхнюю челюсть, скуловую кость (zygomatic bone) и чешуйчатую часть височной кости (tempotal bone) (Рис.7.В).
Вторая жаберная дуга часто называется гиоидной (hyoid arch), так как вносит вклад в образование подъязычной кости. Остальные жаберные дуги обозначаются только номерами.
Мезенхима каждой бранхиальной дуги дает начало мышцам, хрящам и кости.
Клетки нервного гребня дают специфические скелетные структуры, например, эти клетки первой жаберной дуги дают Меккелев хрящ, верхнюю и нижнюю челюсть, различные связки и 2 из 3-х малых косточек, оссиклей, среднего уха. Типичная бранхиальная дуга содержит артерию (aortic arch), полоску хряща, мышечный компонент и нерв. (Рис.5.Б, Г-Е).
Кровь, проходящая через аортальные дуги вступает в дорзальную аорту (Рис.6.Д,Г).
Рис. 8. Рис. 8. А - область головы и шеи эмбриона человека (28 дней). Показаны зачатки мышц бранхиальных дуг. Стрелка показыват путь миграции миобластов из окципитальных миотомов для образования мускулатуры языка. Б - голова и шея плода человека ( 20 недель). Показаны мышцы, происходящие из бранхиальных дуг ( часть мышц platisma и sternocleidomastoid удалена, чтобы показать более глубокие мышцы). Миобласты из 2-й бранхиальной дуги мигрируют в область шеи и дают мышцы лица
1 - затылочные миотомы, 2 - m. orbicularis, 3 - m. buccinator, 4 - m. orbitalis oculi, 5 - m. frontalis, 6 - m. temporalis, 7 - v. occipitalis, 8 - m. auricularis, 9 - m. stylohyoid, 10 - m. stylopharingeus, 11 - anterior and posterior bellies of digastric, 12 - m. platisma, 13 - ключица, 14 - m. sternocleidomastoid, 15 - pharingeal muscles, 16 - m. mylohyoid, 17 - m. masseter
Хрящи бранхиальных дуг. Хрящевой компонент 1-й жаберной дуги называется Меккелевым хрящем (Meckel's cartilage), 2-й дуги - хрящем Рейхерта (Reichert's cartilage). Хрящи 3-й и 4-й дуг не имеют специального названия (см.29-й день, Рис.6). На рис.2 показано положение мышечных компонентов жаберных дуг (Рис.7.А) и их производных на голове и шее 20-недельного плода (Рис.7.Б).
А - вид сбоку на область головы и шеи у эмбриона человека 4-х недель. Показаны черепно-лицевые нервы, производные соответствующих бранхиальных дуг. Б - голова плода (20 недель). Показано поверхностное распределение двух каудальных ветвей тройничного нерва (V), производных первой бранхиальной дуги. В - сагитальный разрез через голову плода. Показано распределение глубоких сенсорных волокон бранхъиальных нервов к зубам, слизистой языка, глотке, полости носа, нёбу и гортани
1 - язык, 2 - нёбо, 3 - полость носа, 4 - паратидная железа, 5 - глотка, 6 - пищевод, 7 - гортань, 8 - низнечелюстная железа, 9 - подъязычные железы.
Состав внеклеточного матрикса
Компонент Хорда Аксиальная
мезенхима
Kлетки Оболочка 0 - отсутствие, (+) - слабая, + - сильная, ++ - очень сильная реакция
* - экспрессия, по-видимому, ограничена базальной мембраной Ходроитин/
дерматан сульфат 0 ++ +
Кератан сульфат 0 0 0
Гиалуроновая кислота 0 + +
Аггрекан 0 0 0
Бигликан + 0 0
Коллаген тип I (+), + (+) + <
Коллаген тип II 0 + 0
Декорин 0 ++ +
Фибронектин 0 ++ (+)
Ламинин (+) +* (+)
Тенасцин 0 + 0
TGF-β + > 0 0
Иммунореактивность и интенсивность окрашивания на 'TGF-beta увеличивается
Протеогликан Декорин обычно взаимодействует с коллагеновыми фибриллами, т,е.
участвует в фибриллогенезе, роль бигликана неизвестна. Коллаген тип II,по-видимому,
участвует в эпителиально-мезенхимных взаимодействиях (оболочка). Гликопротеин
фибронектин, по-видимому, связан с базальной мембраной и может выполнять
индуктивную роль. Молекулы ламинина, по-видимому, локализуются в
эндоплазматическом ретикулеме или аппарате Гольджи. Появление тенасцина в
оболочке хорды, по-видимому, связано с хондрогенезом зачатков тел позвонков и
межпозвоночных дисков, испытывающих преходящую хрящевую дифференцировку.
Секреция почти всех компонентов ВКМ в хорде человека регулируется TGF-beta. Так
TGF-beta индуцирует синтез декорина, который может связываться с фактором роста
через его core белок, модулируя его активность. Сходный механизм негативной петли
обратной связи обнаружен для ламинина.
В аксиальной мезенхиме межклеточные пространства заполнены
хондроитин/дерматан сульфатом и гиалуроновой кислотой. Они, особенно последняя,
ответственны за формирование гидратированных пространств, способбствующих
экспансии ВКМ ,миграции клеток и ингибированию дифференцировки или
хондрогенеза. След., склеротомные клетки иммигрирующие в аксиальную область
находят здесь идеальные условия для миграции и нахождения финальной позиции.
Этому способствует и фибронектин ВКМ аксиальной мезенхимы(Gotz et al., 1995).
Эмбрионы кур (Ст.) Эмбрионы мыши (Ст.)По Theiler's (1989) 16 стадия развития ,10-й день
В кровяных островках,образующихся на ранней сомитной стадии (Е8.0) ангиобласты все еще обнаруживают высокий уровень экспрессии flt-1, тогда как большинство гематопоэтических клеток экспрессирует flt-1
на низком уровне. Далее экспрессия flt-1 в гематопоэтических клетках снижается и наконец, экспрессия ограничивается только эндотелиальными клетками , достигая максимума к среднесомитной стадии (Е8.5). Это указывает на то, что гематопоэтические и эндотелиальные клетки происходят из общего предшественника. Затем экспрессия flt-1 в эндотелиальных клетках снижается и поддерживается на очень низком уровне на поздних стадиях эмбриогенеза.
Образование эмбриональных складок Заканчивается образование головной складки. Зачаток сердца и перикардиальная полость оказываются внутри эмбриона (Рис 3.Б). Хвостовая скадка, возникшая вначале в результате роста спинного мозга в каудальном направлении углубляется в кранио-дорсальном направлении в результате каудальная часть эмбриона оказывается над клоачной мембраной (Рис.1) В это же время часть желточного мешка включается в эмбрион как задняя кишка (hindgut) (зачаток нисходящего колона). Терминальная часть задней кишки слегка расширяется и формирует клоаку ( зачаток мочевого пузыря и прямой кишки). После окончания образования хвостовой складки остаток примитивной полоски оказывается теперь расположенным не краниальнее, а каудальнее клоачной мембраны.
Рис. 5. Эмбрион человека, 28-й день развития. А - внешний вид эмбриона сбоку, амнион удален. Б - сагитальный разрез, показаны составляющие пупочного канатика. В - поперечный разрез по штриховой линии, показанной на схеме Б.
1 - сомит, 2 - пупочный канатик, 3 - орофарингеальная мембрана, 4 - перикардиальная полость, 5 - сердце, 6 - нервная трубка, 7 - хорда, 8 - остатки примитивной полоски, 9 - клоачная мембрана, 10 - ганглии задних корешков спинальных нервов, 11 - внутриэмбриональный целом, 12 - средняя кишка, 13 - вентральная стенка живота, 14 - латеральная стенка живота, 15 - аорта, 16 - урогенитальный гребень Латеральные складки смыкаются по медиальной плоскости, за исключением уровня желточного стебля, устье которого сильно сужается. Область присоединения амниона к эмбриону в целом также сильно сужается , превращаясь в пупочную область (umbilical cord) (Рис.4.Б). Смыкаясь латеральные складки уменьшают область сообщения внутриэмбрионального целома с внеэмбриональным целомом до узкого канала, который сохраняется вплоть до конца 10-й недели (длинная стрелка на рис.2Б).
Примитивная полоска
Примитивная полоска заканчивает активное формировние внутриэмбриональной мезодермы.
Хорда
Хорда представлена непрерывным цилиндром клеток, идущим через развивающийся позвоночный столб. Плотно упакованные клетки хорды содержат значительные отложения гликогена и цитофиламент. Клетки связаны многочисленными десмосомами . Аморфная мембрана вокруг хорды утолщается начинает походить на оболочку,состоящую из тоноко филаментозного материала. Плотность клеток в аксиальной мезенхиме увеличивается.
Брыжейки В результате образования латеральных эмбриональных складок и движения их навстречу друг другу ножки или латеральные части внутриэмбрионального целома также смещаются навстеречу друг другу и в вентральной части эмбриона они встречаются, разделенные дорзальнее и вентральнее сердца и кишки только двумя слоями спланхнической мезодермы (мезокардием и мезентерием) (Рис.4 В). В задней части эмбриона вентральный мезентерий начинает дегенерировать, в результате здесь образуется большая эмбриональная перитонеальная полость. Становятся хорошо различимы три целомические полости: (1) большая перикардиальная полость (результат дегенерации вентрального мезокардия), (2) два относительно небольших перикардиоперитонеальных канала (pericardioperitoneal canals) и (3) большая перитонеальная полость ( результат дегенерации вентрального мезентерия).
Мезентерий желудка.
На данной стадии развития желудок расположен в срединной плоскости и связан с дорзальной стенкой брюшной полости дорзальным мезентерием (dorsal mesentery) часто называемым дорзальным мезогастрием (dorsal mesogastrium), а с вентральной брюшной стенкой он связан вентральным мезентерием или вентральным мезогастрием (Рис. 4 )
Экспансия амниотической полости ведет к облитерации внеэмбриональнго целома и амнинон начинает формировать эпителиальную оболочку для пупочного канатика.
Продолжается процесс сомитогенеза.
Рис. 6 г. Поперечные срезы через эмбрион человека. А - пресомитный эмбрион (около 18 дней),видна параксиальная мезодерма, из которой происходят сомиты. Б - эмбрион около 22 дней. В - эмбрион около 26 дней. Область дермомиотома сомита дает миотом и дерматом (будущая дерма), Г - эмбрион около 28 дней. Стрелками показана миграция клеток из области склеротома сомита
1 - нервная борозда, 2 - нервная пластинка, 3 - латеральная мезодерма, 4 - внеэмбриональный целом, 5 - промежуточная мезодерма, 6 - хорда, 7 - параксиальная мезодерма, 8 - внутриэмбриональный целом, 9 - амнион, 10 - нервная складка, 11 - соматическая мезодерма, 12 - спланхническая мезодерма, 13 - сомит, 14 - амниотическая полость, 15 - нервная трубка, 16 - дермомиотом, 17 - склеротом, 18 - желточный мешок, 19 - дорсальная аорта, 20 - дерматом, 21 - миотом, 22 - первичная кишка
Клетки нервного гребеняКлетки нервного гребня дают специфические скелетные структуры, например, эти клетки первой жаберной дуги дают Меккелев хрящ, верхнюю и нижнюю челюсть, различные связки и 2 из 3-х малых косточек, оссиклей, среднего уха. Типичная бранхиальная дуга содержит артерию (aortic arch), полоску хряща, мышечный компонент и нерв. (Рис.6.Б, Г-Е).
Жаберный аппарат Развитие жаберных дуг начинается с миграции клеток нервного гребня (neural crest cells) в область будущей головы и шеи. В результате проникновения этих клеток образуется незначительное возвышение по бокам развивающейся глотки, первая бранхиальная дуга, зачаток челюстей. На 24-й день развития обнаруживается появление второй жаберной дуги , на 26-й день - третьей и на 28-й день - четвертой жаберной дуги. Ч-я и 6-я жаберные дуги малы и не могут быть видны на поверхности эмбриона. Жаберные дуги отделяются друг от друга хорошо заметными щелями, называемыми жаберными щелями (branchial grooves) (Рис.5А)
Рис. 7. Эмбрион человека (28 дней). Бранхиальный аппарат. Штриховыми линиями обозначены уровни разрезов
1 - , 2 - , 3 - , 4 - 1-4-я бранхиальные дуги, 5 - зачаток языка, 6 - Меккелев хрящ, 7 - первая дуга аорты, 8 - нерв, 9 - слепое отверстие, 10 - пищевод, 11 - ларинготрахеальная борозда, 12 - положение орофарингеальной мембраны, 13 - передний мозг, 14 - верхнечелюстное выпячивание, 15 - хорда, 16 - 1-й глоточный карман, 17 - сердце, 18 - рот, 19 - 3-я дуга аорты, 20 - дорсальная аорта, 21 - зачаток легкого, 22 - truncus arteriosus, 23 - 1-я бранхиальная мембрана, 24, 25 - 2-й и 3-й глоточные карманы, 26 - хрящ Рейхарта, 27 - мышцы, 28 - тироидный дивертикул
Первая жаберная дуга часто называется нижнечелюстной дугой (mandibular arch), она состоит из двух выпячиваний ("отростков") : (1) большого нижнечелюстного выпячивания (mandibular prominence), формирующего нижнюю челюсть, и (2) небольшого верхнечелюстного выпячивания ( maxillary prominence), формирующего верхнюю челюсть, скуловую кость (zygomatic bone) и чешуйчатую часть височной кости (tempotal bone) (Рис.7.В).
Вторая жаберная дуга часто называется гиоидной (hyoid arch), так как вносит вклад в образование подъязычной кости. Остальные жаберные дуги обозначаются только номерами.
Мезенхима каждой бранхиальной дуги дает начало мышцам, хрящам и кости.
Клетки нервного гребня дают специфические скелетные структуры, например, эти клетки первой жаберной дуги дают Меккелев хрящ, верхнюю и нижнюю челюсть, различные связки и 2 из 3-х малых косточек, оссиклей, среднего уха. Типичная бранхиальная дуга содержит артерию (aortic arch), полоску хряща, мышечный компонент и нерв. (Рис.5.Б, Г-Е).
Кровь, проходящая через аортальные дуги вступает в дорзальную аорту (Рис.6.Д,Г).
Рис. 8. Рис. 8. А - область головы и шеи эмбриона человека (28 дней). Показаны зачатки мышц бранхиальных дуг. Стрелка показыват путь миграции миобластов из окципитальных миотомов для образования мускулатуры языка. Б - голова и шея плода человека ( 20 недель). Показаны мышцы, происходящие из бранхиальных дуг ( часть мышц platisma и sternocleidomastoid удалена, чтобы показать более глубокие мышцы). Миобласты из 2-й бранхиальной дуги мигрируют в область шеи и дают мышцы лица
1 - затылочные миотомы, 2 - m. orbicularis, 3 - m. buccinator, 4 - m. orbitalis oculi, 5 - m. frontalis, 6 - m. temporalis, 7 - v. occipitalis, 8 - m. auricularis, 9 - m. stylohyoid, 10 - m. stylopharingeus, 11 - anterior and posterior bellies of digastric, 12 - m. platisma, 13 - ключица, 14 - m. sternocleidomastoid, 15 - pharingeal muscles, 16 - m. mylohyoid, 17 - m. masseter
Хрящи бранхиальных дуг. Хрящевой компонент 1-й жаберной дуги называется Меккелевым хрящем (Meckel's cartilage), 2-й дуги - хрящем Рейхерта (Reichert's cartilage). Хрящи 3-й и 4-й дуг не имеют специального названия (см.29-й день, Рис.6). На рис.2 показано положение мышечных компонентов жаберных дуг (Рис.7.А) и их производных на голове и шее 20-недельного плода (Рис.7.Б).
А - вид сбоку на область головы и шеи у эмбриона человека 4-х недель. Показаны черепно-лицевые нервы, производные соответствующих бранхиальных дуг. Б - голова плода (20 недель). Показано поверхностное распределение двух каудальных ветвей тройничного нерва (V), производных первой бранхиальной дуги. В - сагитальный разрез через голову плода. Показано распределение глубоких сенсорных волокон бранхъиальных нервов к зубам, слизистой языка, глотке, полости носа, нёбу и гортани
1 - язык, 2 - нёбо, 3 - полость носа, 4 - паратидная железа, 5 - глотка, 6 - пищевод, 7 - гортань, 8 - низнечелюстная железа, 9 - подъязычные железы.
Состав внеклеточного матрикса
Компонент Хорда Аксиальная
мезенхима
Kлетки Оболочка 0 - отсутствие, (+) - слабая, + - сильная, ++ - очень сильная реакция
* - экспрессия, по-видимому, ограничена базальной мембраной Ходроитин/
дерматан сульфат 0 ++ +
Кератан сульфат 0 0 0
Гиалуроновая кислота 0 + +
Аггрекан 0 0 0
Бигликан + 0 0
Коллаген тип I (+), + (+) + <
Коллаген тип II 0 + 0
Декорин 0 ++ +
Фибронектин 0 ++ (+)
Ламинин (+) +* (+)
Тенасцин 0 + 0
TGF-β + > 0 0
Иммунореактивность и интенсивность окрашивания на 'TGF-beta увеличивается
Протеогликан Декорин обычно взаимодействует с коллагеновыми фибриллами, т,е.
участвует в фибриллогенезе, роль бигликана неизвестна. Коллаген тип II,по-видимому,
участвует в эпителиально-мезенхимных взаимодействиях (оболочка). Гликопротеин
фибронектин, по-видимому, связан с базальной мембраной и может выполнять
индуктивную роль. Молекулы ламинина, по-видимому, локализуются в
эндоплазматическом ретикулеме или аппарате Гольджи. Появление тенасцина в
оболочке хорды, по-видимому, связано с хондрогенезом зачатков тел позвонков и
межпозвоночных дисков, испытывающих преходящую хрящевую дифференцировку.
Секреция почти всех компонентов ВКМ в хорде человека регулируется TGF-beta. Так
TGF-beta индуцирует синтез декорина, который может связываться с фактором роста
через его core белок, модулируя его активность. Сходный механизм негативной петли
обратной связи обнаружен для ламинина.
В аксиальной мезенхиме межклеточные пространства заполнены
хондроитин/дерматан сульфатом и гиалуроновой кислотой. Они, особенно последняя,
ответственны за формирование гидратированных пространств, способбствующих
экспансии ВКМ ,миграции клеток и ингибированию дифференцировки или
хондрогенеза. След., склеротомные клетки иммигрирующие в аксиальную область
находят здесь идеальные условия для миграции и нахождения финальной позиции.
Этому способствует и фибронектин ВКМ аксиальной мезенхимы(Gotz et al., 1995).
Эмбрионы кур (Ст.) Эмбрионы мыши (Ст.)По Theiler's (1989) 16 стадия развития ,10-й день
В кровяных островках,образующихся на ранней сомитной стадии (Е8.0) ангиобласты все еще обнаруживают высокий уровень экспрессии flt-1, тогда как большинство гематопоэтических клеток экспрессирует flt-1
на низком уровне. Далее экспрессия flt-1 в гематопоэтических клетках снижается и наконец, экспрессия ограничивается только эндотелиальными клетками , достигая максимума к среднесомитной стадии (Е8.5). Это указывает на то, что гематопоэтические и эндотелиальные клетки происходят из общего предшественника. Затем экспрессия flt-1 в эндотелиальных клетках снижается и поддерживается на очень низком уровне на поздних стадиях эмбриогенеза.
1 - сомит, 2 - пупочный канатик, 3 - орофарингеальная мембрана, 4 - перикардиальная полость, 5 - сердце, 6 - нервная трубка, 7 - хорда, 8 - остатки примитивной полоски, 9 - клоачная мембрана, 10 - ганглии задних корешков спинальных нервов, 11 - внутриэмбриональный целом, 12 - средняя кишка, 13 - вентральная стенка живота, 14 - латеральная стенка живота, 15 - аорта, 16 - урогенитальный гребень
Латеральные складки смыкаются по медиальной плоскости, за исключением уровня желточного стебля, устье которого сильно сужается. Область присоединения амниона к эмбриону в целом также сильно сужается , превращаясь в пупочную область (umbilical cord) (Рис.4.Б). Смыкаясь латеральные складки уменьшают область сообщения внутриэмбрионального целома с внеэмбриональным целомом до узкого канала, который сохраняется вплоть до конца 10-й недели (длинная стрелка на рис.2Б).
Примитивная полоска
Примитивная полоска заканчивает активное формировние внутриэмбриональной мезодермы.
Хорда
Хорда представлена непрерывным цилиндром клеток, идущим через развивающийся позвоночный столб. Плотно упакованные клетки хорды содержат значительные отложения гликогена и цитофиламент. Клетки связаны многочисленными десмосомами . Аморфная мембрана вокруг хорды утолщается начинает походить на оболочку,состоящую из тоноко филаментозного материала. Плотность клеток в аксиальной мезенхиме увеличивается.
Брыжейки В результате образования латеральных эмбриональных складок и движения их навстречу друг другу ножки или латеральные части внутриэмбрионального целома также смещаются навстеречу друг другу и в вентральной части эмбриона они встречаются, разделенные дорзальнее и вентральнее сердца и кишки только двумя слоями спланхнической мезодермы (мезокардием и мезентерием) (Рис.4 В). В задней части эмбриона вентральный мезентерий начинает дегенерировать, в результате здесь образуется большая эмбриональная перитонеальная полость. Становятся хорошо различимы три целомические полости: (1) большая перикардиальная полость (результат дегенерации вентрального мезокардия), (2) два относительно небольших перикардиоперитонеальных канала (pericardioperitoneal canals) и (3) большая перитонеальная полость ( результат дегенерации вентрального мезентерия).
Мезентерий желудка.
На данной стадии развития желудок расположен в срединной плоскости и связан с дорзальной стенкой брюшной полости дорзальным мезентерием (dorsal mesentery) часто называемым дорзальным мезогастрием (dorsal mesogastrium), а с вентральной брюшной стенкой он связан вентральным мезентерием или вентральным мезогастрием (Рис. 4 )
Экспансия амниотической полости ведет к облитерации внеэмбриональнго целома и амнинон начинает формировать эпителиальную оболочку для пупочного канатика.
Продолжается процесс сомитогенеза.
Рис. 6 г. Поперечные срезы через эмбрион человека. А - пресомитный эмбрион (около 18 дней),видна параксиальная мезодерма, из которой происходят сомиты. Б - эмбрион около 22 дней. В - эмбрион около 26 дней. Область дермомиотома сомита дает миотом и дерматом (будущая дерма), Г - эмбрион около 28 дней. Стрелками показана миграция клеток из области склеротома сомита
1 - нервная борозда, 2 - нервная пластинка, 3 - латеральная мезодерма, 4 - внеэмбриональный целом, 5 - промежуточная мезодерма, 6 - хорда, 7 - параксиальная мезодерма, 8 - внутриэмбриональный целом, 9 - амнион, 10 - нервная складка, 11 - соматическая мезодерма, 12 - спланхническая мезодерма, 13 - сомит, 14 - амниотическая полость, 15 - нервная трубка, 16 - дермомиотом, 17 - склеротом, 18 - желточный мешок, 19 - дорсальная аорта, 20 - дерматом, 21 - миотом, 22 - первичная кишка
Клетки нервного гребеняКлетки нервного гребня дают специфические скелетные структуры, например, эти клетки первой жаберной дуги дают Меккелев хрящ, верхнюю и нижнюю челюсть, различные связки и 2 из 3-х малых косточек, оссиклей, среднего уха. Типичная бранхиальная дуга содержит артерию (aortic arch), полоску хряща, мышечный компонент и нерв. (Рис.6.Б, Г-Е).
Жаберный аппарат Развитие жаберных дуг начинается с миграции клеток нервного гребня (neural crest cells) в область будущей головы и шеи. В результате проникновения этих клеток образуется незначительное возвышение по бокам развивающейся глотки, первая бранхиальная дуга, зачаток челюстей. На 24-й день развития обнаруживается появление второй жаберной дуги , на 26-й день - третьей и на 28-й день - четвертой жаберной дуги. Ч-я и 6-я жаберные дуги малы и не могут быть видны на поверхности эмбриона. Жаберные дуги отделяются друг от друга хорошо заметными щелями, называемыми жаберными щелями (branchial grooves) (Рис.5А)
Рис. 7. Эмбрион человека (28 дней). Бранхиальный аппарат. Штриховыми линиями обозначены уровни разрезов
1 - , 2 - , 3 - , 4 - 1-4-я бранхиальные дуги, 5 - зачаток языка, 6 - Меккелев хрящ, 7 - первая дуга аорты, 8 - нерв, 9 - слепое отверстие, 10 - пищевод, 11 - ларинготрахеальная борозда, 12 - положение орофарингеальной мембраны, 13 - передний мозг, 14 - верхнечелюстное выпячивание, 15 - хорда, 16 - 1-й глоточный карман, 17 - сердце, 18 - рот, 19 - 3-я дуга аорты, 20 - дорсальная аорта, 21 - зачаток легкого, 22 - truncus arteriosus, 23 - 1-я бранхиальная мембрана, 24, 25 - 2-й и 3-й глоточные карманы, 26 - хрящ Рейхарта, 27 - мышцы, 28 - тироидный дивертикул
Первая жаберная дуга часто называется нижнечелюстной дугой (mandibular arch), она состоит из двух выпячиваний ("отростков") : (1) большого нижнечелюстного выпячивания (mandibular prominence), формирующего нижнюю челюсть, и (2) небольшого верхнечелюстного выпячивания ( maxillary prominence), формирующего верхнюю челюсть, скуловую кость (zygomatic bone) и чешуйчатую часть височной кости (tempotal bone) (Рис.7.В).
Вторая жаберная дуга часто называется гиоидной (hyoid arch), так как вносит вклад в образование подъязычной кости. Остальные жаберные дуги обозначаются только номерами.
Мезенхима каждой бранхиальной дуги дает начало мышцам, хрящам и кости.
Клетки нервного гребня дают специфические скелетные структуры, например, эти клетки первой жаберной дуги дают Меккелев хрящ, верхнюю и нижнюю челюсть, различные связки и 2 из 3-х малых косточек, оссиклей, среднего уха. Типичная бранхиальная дуга содержит артерию (aortic arch), полоску хряща, мышечный компонент и нерв. (Рис.5.Б, Г-Е).
Кровь, проходящая через аортальные дуги вступает в дорзальную аорту (Рис.6.Д,Г).
Рис. 8. Рис. 8. А - область головы и шеи эмбриона человека (28 дней). Показаны зачатки мышц бранхиальных дуг. Стрелка показыват путь миграции миобластов из окципитальных миотомов для образования мускулатуры языка. Б - голова и шея плода человека ( 20 недель). Показаны мышцы, происходящие из бранхиальных дуг ( часть мышц platisma и sternocleidomastoid удалена, чтобы показать более глубокие мышцы). Миобласты из 2-й бранхиальной дуги мигрируют в область шеи и дают мышцы лица
1 - затылочные миотомы, 2 - m. orbicularis, 3 - m. buccinator, 4 - m. orbitalis oculi, 5 - m. frontalis, 6 - m. temporalis, 7 - v. occipitalis, 8 - m. auricularis, 9 - m. stylohyoid, 10 - m. stylopharingeus, 11 - anterior and posterior bellies of digastric, 12 - m. platisma, 13 - ключица, 14 - m. sternocleidomastoid, 15 - pharingeal muscles, 16 - m. mylohyoid, 17 - m. masseter
Хрящи бранхиальных дуг. Хрящевой компонент 1-й жаберной дуги называется Меккелевым хрящем (Meckel's cartilage), 2-й дуги - хрящем Рейхерта (Reichert's cartilage). Хрящи 3-й и 4-й дуг не имеют специального названия (см.29-й день, Рис.6). На рис.2 показано положение мышечных компонентов жаберных дуг (Рис.7.А) и их производных на голове и шее 20-недельного плода (Рис.7.Б).
А - вид сбоку на область головы и шеи у эмбриона человека 4-х недель. Показаны черепно-лицевые нервы, производные соответствующих бранхиальных дуг. Б - голова плода (20 недель). Показано поверхностное распределение двух каудальных ветвей тройничного нерва (V), производных первой бранхиальной дуги. В - сагитальный разрез через голову плода. Показано распределение глубоких сенсорных волокон бранхъиальных нервов к зубам, слизистой языка, глотке, полости носа, нёбу и гортани
1 - язык, 2 - нёбо, 3 - полость носа, 4 - паратидная железа, 5 - глотка, 6 - пищевод, 7 - гортань, 8 - низнечелюстная железа, 9 - подъязычные железы.
Состав внеклеточного матрикса
Компонент Хорда Аксиальная
мезенхима
Kлетки Оболочка 0 - отсутствие, (+) - слабая, + - сильная, ++ - очень сильная реакция
* - экспрессия, по-видимому, ограничена базальной мембраной Ходроитин/
дерматан сульфат 0 ++ +
Кератан сульфат 0 0 0
Гиалуроновая кислота 0 + +
Аггрекан 0 0 0
Бигликан + 0 0
Коллаген тип I (+), + (+) + <
Коллаген тип II 0 + 0
Декорин 0 ++ +
Фибронектин 0 ++ (+)
Ламинин (+) +* (+)
Тенасцин 0 + 0
TGF-β + > 0 0
Иммунореактивность и интенсивность окрашивания на 'TGF-beta увеличивается
Протеогликан Декорин обычно взаимодействует с коллагеновыми фибриллами, т,е.
участвует в фибриллогенезе, роль бигликана неизвестна. Коллаген тип II,по-видимому,
участвует в эпителиально-мезенхимных взаимодействиях (оболочка). Гликопротеин
фибронектин, по-видимому, связан с базальной мембраной и может выполнять
индуктивную роль. Молекулы ламинина, по-видимому, локализуются в
эндоплазматическом ретикулеме или аппарате Гольджи. Появление тенасцина в
оболочке хорды, по-видимому, связано с хондрогенезом зачатков тел позвонков и
межпозвоночных дисков, испытывающих преходящую хрящевую дифференцировку.
Секреция почти всех компонентов ВКМ в хорде человека регулируется TGF-beta. Так
TGF-beta индуцирует синтез декорина, который может связываться с фактором роста
через его core белок, модулируя его активность. Сходный механизм негативной петли
обратной связи обнаружен для ламинина.
В аксиальной мезенхиме межклеточные пространства заполнены
хондроитин/дерматан сульфатом и гиалуроновой кислотой. Они, особенно последняя,
ответственны за формирование гидратированных пространств, способбствующих
экспансии ВКМ ,миграции клеток и ингибированию дифференцировки или
хондрогенеза. След., склеротомные клетки иммигрирующие в аксиальную область
находят здесь идеальные условия для миграции и нахождения финальной позиции.
Этому способствует и фибронектин ВКМ аксиальной мезенхимы(Gotz et al., 1995).
Эмбрионы кур (Ст.) Эмбрионы мыши (Ст.)По Theiler's (1989) 16 стадия развития ,10-й день
В кровяных островках,образующихся на ранней сомитной стадии (Е8.0) ангиобласты все еще обнаруживают высокий уровень экспрессии flt-1, тогда как большинство гематопоэтических клеток экспрессирует flt-1
на низком уровне. Далее экспрессия flt-1 в гематопоэтических клетках снижается и наконец, экспрессия ограничивается только эндотелиальными клетками , достигая максимума к среднесомитной стадии (Е8.5). Это указывает на то, что гематопоэтические и эндотелиальные клетки происходят из общего предшественника. Затем экспрессия flt-1 в эндотелиальных клетках снижается и поддерживается на очень низком уровне на поздних стадиях эмбриогенеза.
Хорда
Хорда представлена непрерывным цилиндром клеток, идущим через развивающийся позвоночный столб. Плотно упакованные клетки хорды содержат значительные отложения гликогена и цитофиламент. Клетки связаны многочисленными десмосомами . Аморфная мембрана вокруг хорды утолщается начинает походить на оболочку,состоящую из тоноко филаментозного материала. Плотность клеток в аксиальной мезенхиме увеличивается.
Брыжейки В результате образования латеральных эмбриональных складок и движения их навстречу друг другу ножки или латеральные части внутриэмбрионального целома также смещаются навстеречу друг другу и в вентральной части эмбриона они встречаются, разделенные дорзальнее и вентральнее сердца и кишки только двумя слоями спланхнической мезодермы (мезокардием и мезентерием) (Рис.4 В). В задней части эмбриона вентральный мезентерий начинает дегенерировать, в результате здесь образуется большая эмбриональная перитонеальная полость. Становятся хорошо различимы три целомические полости: (1) большая перикардиальная полость (результат дегенерации вентрального мезокардия), (2) два относительно небольших перикардиоперитонеальных канала (pericardioperitoneal canals) и (3) большая перитонеальная полость ( результат дегенерации вентрального мезентерия).
Мезентерий желудка.
На данной стадии развития желудок расположен в срединной плоскости и связан с дорзальной стенкой брюшной полости дорзальным мезентерием (dorsal mesentery) часто называемым дорзальным мезогастрием (dorsal mesogastrium), а с вентральной брюшной стенкой он связан вентральным мезентерием или вентральным мезогастрием (Рис. 4 )
Экспансия амниотической полости ведет к облитерации внеэмбриональнго целома и амнинон начинает формировать эпителиальную оболочку для пупочного канатика.
Продолжается процесс сомитогенеза.
Рис. 6 г. Поперечные срезы через эмбрион человека. А - пресомитный эмбрион (около 18 дней),видна параксиальная мезодерма, из которой происходят сомиты. Б - эмбрион около 22 дней. В - эмбрион около 26 дней. Область дермомиотома сомита дает миотом и дерматом (будущая дерма), Г - эмбрион около 28 дней. Стрелками показана миграция клеток из области склеротома сомита
1 - нервная борозда, 2 - нервная пластинка, 3 - латеральная мезодерма, 4 - внеэмбриональный целом, 5 - промежуточная мезодерма, 6 - хорда, 7 - параксиальная мезодерма, 8 - внутриэмбриональный целом, 9 - амнион, 10 - нервная складка, 11 - соматическая мезодерма, 12 - спланхническая мезодерма, 13 - сомит, 14 - амниотическая полость, 15 - нервная трубка, 16 - дермомиотом, 17 - склеротом, 18 - желточный мешок, 19 - дорсальная аорта, 20 - дерматом, 21 - миотом, 22 - первичная кишка
Клетки нервного гребеняКлетки нервного гребня дают специфические скелетные структуры, например, эти клетки первой жаберной дуги дают Меккелев хрящ, верхнюю и нижнюю челюсть, различные связки и 2 из 3-х малых косточек, оссиклей, среднего уха. Типичная бранхиальная дуга содержит артерию (aortic arch), полоску хряща, мышечный компонент и нерв. (Рис.6.Б, Г-Е).
Жаберный аппарат Развитие жаберных дуг начинается с миграции клеток нервного гребня (neural crest cells) в область будущей головы и шеи. В результате проникновения этих клеток образуется незначительное возвышение по бокам развивающейся глотки, первая бранхиальная дуга, зачаток челюстей. На 24-й день развития обнаруживается появление второй жаберной дуги , на 26-й день - третьей и на 28-й день - четвертой жаберной дуги. Ч-я и 6-я жаберные дуги малы и не могут быть видны на поверхности эмбриона. Жаберные дуги отделяются друг от друга хорошо заметными щелями, называемыми жаберными щелями (branchial grooves) (Рис.5А)
Рис. 7. Эмбрион человека (28 дней). Бранхиальный аппарат. Штриховыми линиями обозначены уровни разрезов
1 - , 2 - , 3 - , 4 - 1-4-я бранхиальные дуги, 5 - зачаток языка, 6 - Меккелев хрящ, 7 - первая дуга аорты, 8 - нерв, 9 - слепое отверстие, 10 - пищевод, 11 - ларинготрахеальная борозда, 12 - положение орофарингеальной мембраны, 13 - передний мозг, 14 - верхнечелюстное выпячивание, 15 - хорда, 16 - 1-й глоточный карман, 17 - сердце, 18 - рот, 19 - 3-я дуга аорты, 20 - дорсальная аорта, 21 - зачаток легкого, 22 - truncus arteriosus, 23 - 1-я бранхиальная мембрана, 24, 25 - 2-й и 3-й глоточные карманы, 26 - хрящ Рейхарта, 27 - мышцы, 28 - тироидный дивертикул
Первая жаберная дуга часто называется нижнечелюстной дугой (mandibular arch), она состоит из двух выпячиваний ("отростков") : (1) большого нижнечелюстного выпячивания (mandibular prominence), формирующего нижнюю челюсть, и (2) небольшого верхнечелюстного выпячивания ( maxillary prominence), формирующего верхнюю челюсть, скуловую кость (zygomatic bone) и чешуйчатую часть височной кости (tempotal bone) (Рис.7.В).
Вторая жаберная дуга часто называется гиоидной (hyoid arch), так как вносит вклад в образование подъязычной кости. Остальные жаберные дуги обозначаются только номерами.
Мезенхима каждой бранхиальной дуги дает начало мышцам, хрящам и кости.
Клетки нервного гребня дают специфические скелетные структуры, например, эти клетки первой жаберной дуги дают Меккелев хрящ, верхнюю и нижнюю челюсть, различные связки и 2 из 3-х малых косточек, оссиклей, среднего уха. Типичная бранхиальная дуга содержит артерию (aortic arch), полоску хряща, мышечный компонент и нерв. (Рис.5.Б, Г-Е).
Кровь, проходящая через аортальные дуги вступает в дорзальную аорту (Рис.6.Д,Г).
Рис. 8. Рис. 8. А - область головы и шеи эмбриона человека (28 дней). Показаны зачатки мышц бранхиальных дуг. Стрелка показыват путь миграции миобластов из окципитальных миотомов для образования мускулатуры языка. Б - голова и шея плода человека ( 20 недель). Показаны мышцы, происходящие из бранхиальных дуг ( часть мышц platisma и sternocleidomastoid удалена, чтобы показать более глубокие мышцы). Миобласты из 2-й бранхиальной дуги мигрируют в область шеи и дают мышцы лица
1 - затылочные миотомы, 2 - m. orbicularis, 3 - m. buccinator, 4 - m. orbitalis oculi, 5 - m. frontalis, 6 - m. temporalis, 7 - v. occipitalis, 8 - m. auricularis, 9 - m. stylohyoid, 10 - m. stylopharingeus, 11 - anterior and posterior bellies of digastric, 12 - m. platisma, 13 - ключица, 14 - m. sternocleidomastoid, 15 - pharingeal muscles, 16 - m. mylohyoid, 17 - m. masseter
Хрящи бранхиальных дуг. Хрящевой компонент 1-й жаберной дуги называется Меккелевым хрящем (Meckel's cartilage), 2-й дуги - хрящем Рейхерта (Reichert's cartilage). Хрящи 3-й и 4-й дуг не имеют специального названия (см.29-й день, Рис.6). На рис.2 показано положение мышечных компонентов жаберных дуг (Рис.7.А) и их производных на голове и шее 20-недельного плода (Рис.7.Б).
А - вид сбоку на область головы и шеи у эмбриона человека 4-х недель. Показаны черепно-лицевые нервы, производные соответствующих бранхиальных дуг. Б - голова плода (20 недель). Показано поверхностное распределение двух каудальных ветвей тройничного нерва (V), производных первой бранхиальной дуги. В - сагитальный разрез через голову плода. Показано распределение глубоких сенсорных волокон бранхъиальных нервов к зубам, слизистой языка, глотке, полости носа, нёбу и гортани
1 - язык, 2 - нёбо, 3 - полость носа, 4 - паратидная железа, 5 - глотка, 6 - пищевод, 7 - гортань, 8 - низнечелюстная железа, 9 - подъязычные железы.
Состав внеклеточного матрикса
Компонент Хорда Аксиальная
мезенхима
Kлетки Оболочка 0 - отсутствие, (+) - слабая, + - сильная, ++ - очень сильная реакция
* - экспрессия, по-видимому, ограничена базальной мембраной Ходроитин/
дерматан сульфат 0 ++ +
Кератан сульфат 0 0 0
Гиалуроновая кислота 0 + +
Аггрекан 0 0 0
Бигликан + 0 0
Коллаген тип I (+), + (+) + <
Коллаген тип II 0 + 0
Декорин 0 ++ +
Фибронектин 0 ++ (+)
Ламинин (+) +* (+)
Тенасцин 0 + 0
TGF-β + > 0 0
Иммунореактивность и интенсивность окрашивания на 'TGF-beta увеличивается
Протеогликан Декорин обычно взаимодействует с коллагеновыми фибриллами, т,е.
участвует в фибриллогенезе, роль бигликана неизвестна. Коллаген тип II,по-видимому,
участвует в эпителиально-мезенхимных взаимодействиях (оболочка). Гликопротеин
фибронектин, по-видимому, связан с базальной мембраной и может выполнять
индуктивную роль. Молекулы ламинина, по-видимому, локализуются в
эндоплазматическом ретикулеме или аппарате Гольджи. Появление тенасцина в
оболочке хорды, по-видимому, связано с хондрогенезом зачатков тел позвонков и
межпозвоночных дисков, испытывающих преходящую хрящевую дифференцировку.
Секреция почти всех компонентов ВКМ в хорде человека регулируется TGF-beta. Так
TGF-beta индуцирует синтез декорина, который может связываться с фактором роста
через его core белок, модулируя его активность. Сходный механизм негативной петли
обратной связи обнаружен для ламинина.
В аксиальной мезенхиме межклеточные пространства заполнены
хондроитин/дерматан сульфатом и гиалуроновой кислотой. Они, особенно последняя,
ответственны за формирование гидратированных пространств, способбствующих
экспансии ВКМ ,миграции клеток и ингибированию дифференцировки или
хондрогенеза. След., склеротомные клетки иммигрирующие в аксиальную область
находят здесь идеальные условия для миграции и нахождения финальной позиции.
Этому способствует и фибронектин ВКМ аксиальной мезенхимы(Gotz et al., 1995).
Эмбрионы кур (Ст.) Эмбрионы мыши (Ст.)По Theiler's (1989) 16 стадия развития ,10-й день
В кровяных островках,образующихся на ранней сомитной стадии (Е8.0) ангиобласты все еще обнаруживают высокий уровень экспрессии flt-1, тогда как большинство гематопоэтических клеток экспрессирует flt-1
на низком уровне. Далее экспрессия flt-1 в гематопоэтических клетках снижается и наконец, экспрессия ограничивается только эндотелиальными клетками , достигая максимума к среднесомитной стадии (Е8.5). Это указывает на то, что гематопоэтические и эндотелиальные клетки происходят из общего предшественника. Затем экспрессия flt-1 в эндотелиальных клетках снижается и поддерживается на очень низком уровне на поздних стадиях эмбриогенеза.
Брыжейки В результате образования латеральных эмбриональных складок и движения их навстречу друг другу ножки или латеральные части внутриэмбрионального целома также смещаются навстеречу друг другу и в вентральной части эмбриона они встречаются, разделенные дорзальнее и вентральнее сердца и кишки только двумя слоями спланхнической мезодермы (мезокардием и мезентерием) (Рис.4 В). В задней части эмбриона вентральный мезентерий начинает дегенерировать, в результате здесь образуется большая эмбриональная перитонеальная полость. Становятся хорошо различимы три целомические полости: (1) большая перикардиальная полость (результат дегенерации вентрального мезокардия), (2) два относительно небольших перикардиоперитонеальных канала (pericardioperitoneal canals) и (3) большая перитонеальная полость ( результат дегенерации вентрального мезентерия).
Мезентерий желудка.
На данной стадии развития желудок расположен в срединной плоскости и связан с дорзальной стенкой брюшной полости дорзальным мезентерием (dorsal mesentery) часто называемым дорзальным мезогастрием (dorsal mesogastrium), а с вентральной брюшной стенкой он связан вентральным мезентерием или вентральным мезогастрием (Рис. 4 )
Экспансия амниотической полости ведет к облитерации внеэмбриональнго целома и амнинон начинает формировать эпителиальную оболочку для пупочного канатика.
Продолжается процесс сомитогенеза.
Рис. 6 г. Поперечные срезы через эмбрион человека. А - пресомитный эмбрион (около 18 дней),видна параксиальная мезодерма, из которой происходят сомиты. Б - эмбрион около 22 дней. В - эмбрион около 26 дней. Область дермомиотома сомита дает миотом и дерматом (будущая дерма), Г - эмбрион около 28 дней. Стрелками показана миграция клеток из области склеротома сомита
1 - нервная борозда, 2 - нервная пластинка, 3 - латеральная мезодерма, 4 - внеэмбриональный целом, 5 - промежуточная мезодерма, 6 - хорда, 7 - параксиальная мезодерма, 8 - внутриэмбриональный целом, 9 - амнион, 10 - нервная складка, 11 - соматическая мезодерма, 12 - спланхническая мезодерма, 13 - сомит, 14 - амниотическая полость, 15 - нервная трубка, 16 - дермомиотом, 17 - склеротом, 18 - желточный мешок, 19 - дорсальная аорта, 20 - дерматом, 21 - миотом, 22 - первичная кишка
Клетки нервного гребеняКлетки нервного гребня дают специфические скелетные структуры, например, эти клетки первой жаберной дуги дают Меккелев хрящ, верхнюю и нижнюю челюсть, различные связки и 2 из 3-х малых косточек, оссиклей, среднего уха. Типичная бранхиальная дуга содержит артерию (aortic arch), полоску хряща, мышечный компонент и нерв. (Рис.6.Б, Г-Е).
Жаберный аппарат Развитие жаберных дуг начинается с миграции клеток нервного гребня (neural crest cells) в область будущей головы и шеи. В результате проникновения этих клеток образуется незначительное возвышение по бокам развивающейся глотки, первая бранхиальная дуга, зачаток челюстей. На 24-й день развития обнаруживается появление второй жаберной дуги , на 26-й день - третьей и на 28-й день - четвертой жаберной дуги. Ч-я и 6-я жаберные дуги малы и не могут быть видны на поверхности эмбриона. Жаберные дуги отделяются друг от друга хорошо заметными щелями, называемыми жаберными щелями (branchial grooves) (Рис.5А)
Рис. 7. Эмбрион человека (28 дней). Бранхиальный аппарат. Штриховыми линиями обозначены уровни разрезов
1 - , 2 - , 3 - , 4 - 1-4-я бранхиальные дуги, 5 - зачаток языка, 6 - Меккелев хрящ, 7 - первая дуга аорты, 8 - нерв, 9 - слепое отверстие, 10 - пищевод, 11 - ларинготрахеальная борозда, 12 - положение орофарингеальной мембраны, 13 - передний мозг, 14 - верхнечелюстное выпячивание, 15 - хорда, 16 - 1-й глоточный карман, 17 - сердце, 18 - рот, 19 - 3-я дуга аорты, 20 - дорсальная аорта, 21 - зачаток легкого, 22 - truncus arteriosus, 23 - 1-я бранхиальная мембрана, 24, 25 - 2-й и 3-й глоточные карманы, 26 - хрящ Рейхарта, 27 - мышцы, 28 - тироидный дивертикул
Первая жаберная дуга часто называется нижнечелюстной дугой (mandibular arch), она состоит из двух выпячиваний ("отростков") : (1) большого нижнечелюстного выпячивания (mandibular prominence), формирующего нижнюю челюсть, и (2) небольшого верхнечелюстного выпячивания ( maxillary prominence), формирующего верхнюю челюсть, скуловую кость (zygomatic bone) и чешуйчатую часть височной кости (tempotal bone) (Рис.7.В).
Вторая жаберная дуга часто называется гиоидной (hyoid arch), так как вносит вклад в образование подъязычной кости. Остальные жаберные дуги обозначаются только номерами.
Мезенхима каждой бранхиальной дуги дает начало мышцам, хрящам и кости.
Клетки нервного гребня дают специфические скелетные структуры, например, эти клетки первой жаберной дуги дают Меккелев хрящ, верхнюю и нижнюю челюсть, различные связки и 2 из 3-х малых косточек, оссиклей, среднего уха. Типичная бранхиальная дуга содержит артерию (aortic arch), полоску хряща, мышечный компонент и нерв. (Рис.5.Б, Г-Е).
Кровь, проходящая через аортальные дуги вступает в дорзальную аорту (Рис.6.Д,Г).
Рис. 8. Рис. 8. А - область головы и шеи эмбриона человека (28 дней). Показаны зачатки мышц бранхиальных дуг. Стрелка показыват путь миграции миобластов из окципитальных миотомов для образования мускулатуры языка. Б - голова и шея плода человека ( 20 недель). Показаны мышцы, происходящие из бранхиальных дуг ( часть мышц platisma и sternocleidomastoid удалена, чтобы показать более глубокие мышцы). Миобласты из 2-й бранхиальной дуги мигрируют в область шеи и дают мышцы лица
1 - затылочные миотомы, 2 - m. orbicularis, 3 - m. buccinator, 4 - m. orbitalis oculi, 5 - m. frontalis, 6 - m. temporalis, 7 - v. occipitalis, 8 - m. auricularis, 9 - m. stylohyoid, 10 - m. stylopharingeus, 11 - anterior and posterior bellies of digastric, 12 - m. platisma, 13 - ключица, 14 - m. sternocleidomastoid, 15 - pharingeal muscles, 16 - m. mylohyoid, 17 - m. masseter
Хрящи бранхиальных дуг. Хрящевой компонент 1-й жаберной дуги называется Меккелевым хрящем (Meckel's cartilage), 2-й дуги - хрящем Рейхерта (Reichert's cartilage). Хрящи 3-й и 4-й дуг не имеют специального названия (см.29-й день, Рис.6). На рис.2 показано положение мышечных компонентов жаберных дуг (Рис.7.А) и их производных на голове и шее 20-недельного плода (Рис.7.Б).
А - вид сбоку на область головы и шеи у эмбриона человека 4-х недель. Показаны черепно-лицевые нервы, производные соответствующих бранхиальных дуг. Б - голова плода (20 недель). Показано поверхностное распределение двух каудальных ветвей тройничного нерва (V), производных первой бранхиальной дуги. В - сагитальный разрез через голову плода. Показано распределение глубоких сенсорных волокон бранхъиальных нервов к зубам, слизистой языка, глотке, полости носа, нёбу и гортани
1 - язык, 2 - нёбо, 3 - полость носа, 4 - паратидная железа, 5 - глотка, 6 - пищевод, 7 - гортань, 8 - низнечелюстная железа, 9 - подъязычные железы.
Состав внеклеточного матрикса
Компонент Хорда Аксиальная
мезенхима
Kлетки Оболочка 0 - отсутствие, (+) - слабая, + - сильная, ++ - очень сильная реакция
* - экспрессия, по-видимому, ограничена базальной мембраной Ходроитин/
дерматан сульфат 0 ++ +
Кератан сульфат 0 0 0
Гиалуроновая кислота 0 + +
Аггрекан 0 0 0
Бигликан + 0 0
Коллаген тип I (+), + (+) + <
Коллаген тип II 0 + 0
Декорин 0 ++ +
Фибронектин 0 ++ (+)
Ламинин (+) +* (+)
Тенасцин 0 + 0
TGF-β + > 0 0
Иммунореактивность и интенсивность окрашивания на 'TGF-beta увеличивается
Протеогликан Декорин обычно взаимодействует с коллагеновыми фибриллами, т,е.
участвует в фибриллогенезе, роль бигликана неизвестна. Коллаген тип II,по-видимому,
участвует в эпителиально-мезенхимных взаимодействиях (оболочка). Гликопротеин
фибронектин, по-видимому, связан с базальной мембраной и может выполнять
индуктивную роль. Молекулы ламинина, по-видимому, локализуются в
эндоплазматическом ретикулеме или аппарате Гольджи. Появление тенасцина в
оболочке хорды, по-видимому, связано с хондрогенезом зачатков тел позвонков и
межпозвоночных дисков, испытывающих преходящую хрящевую дифференцировку.
Секреция почти всех компонентов ВКМ в хорде человека регулируется TGF-beta. Так
TGF-beta индуцирует синтез декорина, который может связываться с фактором роста
через его core белок, модулируя его активность. Сходный механизм негативной петли
обратной связи обнаружен для ламинина.
В аксиальной мезенхиме межклеточные пространства заполнены
хондроитин/дерматан сульфатом и гиалуроновой кислотой. Они, особенно последняя,
ответственны за формирование гидратированных пространств, способбствующих
экспансии ВКМ ,миграции клеток и ингибированию дифференцировки или
хондрогенеза. След., склеротомные клетки иммигрирующие в аксиальную область
находят здесь идеальные условия для миграции и нахождения финальной позиции.
Этому способствует и фибронектин ВКМ аксиальной мезенхимы(Gotz et al., 1995).
Эмбрионы кур (Ст.) Эмбрионы мыши (Ст.)По Theiler's (1989) 16 стадия развития ,10-й день
В кровяных островках,образующихся на ранней сомитной стадии (Е8.0) ангиобласты все еще обнаруживают высокий уровень экспрессии flt-1, тогда как большинство гематопоэтических клеток экспрессирует flt-1
на низком уровне. Далее экспрессия flt-1 в гематопоэтических клетках снижается и наконец, экспрессия ограничивается только эндотелиальными клетками , достигая максимума к среднесомитной стадии (Е8.5). Это указывает на то, что гематопоэтические и эндотелиальные клетки происходят из общего предшественника. Затем экспрессия flt-1 в эндотелиальных клетках снижается и поддерживается на очень низком уровне на поздних стадиях эмбриогенеза.
Мезентерий желудка.
Продолжается процесс сомитогенеза.
1 - нервная борозда, 2 - нервная пластинка, 3 - латеральная мезодерма, 4 - внеэмбриональный целом, 5 - промежуточная мезодерма, 6 - хорда, 7 - параксиальная мезодерма, 8 - внутриэмбриональный целом, 9 - амнион, 10 - нервная складка, 11 - соматическая мезодерма, 12 - спланхническая мезодерма, 13 - сомит, 14 - амниотическая полость, 15 - нервная трубка, 16 - дермомиотом, 17 - склеротом, 18 - желточный мешок, 19 - дорсальная аорта, 20 - дерматом, 21 - миотом, 22 - первичная кишка
Клетки нервного гребеняКлетки нервного гребня дают специфические скелетные структуры, например, эти клетки первой жаберной дуги дают Меккелев хрящ, верхнюю и нижнюю челюсть, различные связки и 2 из 3-х малых косточек, оссиклей, среднего уха. Типичная бранхиальная дуга содержит артерию (aortic arch), полоску хряща, мышечный компонент и нерв. (Рис.6.Б, Г-Е).
Жаберный аппарат Развитие жаберных дуг начинается с миграции клеток нервного гребня (neural crest cells) в область будущей головы и шеи. В результате проникновения этих клеток образуется незначительное возвышение по бокам развивающейся глотки, первая бранхиальная дуга, зачаток челюстей. На 24-й день развития обнаруживается появление второй жаберной дуги , на 26-й день - третьей и на 28-й день - четвертой жаберной дуги. Ч-я и 6-я жаберные дуги малы и не могут быть видны на поверхности эмбриона. Жаберные дуги отделяются друг от друга хорошо заметными щелями, называемыми жаберными щелями (branchial grooves) (Рис.5А)
Рис. 7. Эмбрион человека (28 дней). Бранхиальный аппарат. Штриховыми линиями обозначены уровни разрезов
1 - , 2 - , 3 - , 4 - 1-4-я бранхиальные дуги, 5 - зачаток языка, 6 - Меккелев хрящ, 7 - первая дуга аорты, 8 - нерв, 9 - слепое отверстие, 10 - пищевод, 11 - ларинготрахеальная борозда, 12 - положение орофарингеальной мембраны, 13 - передний мозг, 14 - верхнечелюстное выпячивание, 15 - хорда, 16 - 1-й глоточный карман, 17 - сердце, 18 - рот, 19 - 3-я дуга аорты, 20 - дорсальная аорта, 21 - зачаток легкого, 22 - truncus arteriosus, 23 - 1-я бранхиальная мембрана, 24, 25 - 2-й и 3-й глоточные карманы, 26 - хрящ Рейхарта, 27 - мышцы, 28 - тироидный дивертикул
Первая жаберная дуга часто называется нижнечелюстной дугой (mandibular arch), она состоит из двух выпячиваний ("отростков") : (1) большого нижнечелюстного выпячивания (mandibular prominence), формирующего нижнюю челюсть, и (2) небольшого верхнечелюстного выпячивания ( maxillary prominence), формирующего верхнюю челюсть, скуловую кость (zygomatic bone) и чешуйчатую часть височной кости (tempotal bone) (Рис.7.В).
Вторая жаберная дуга часто называется гиоидной (hyoid arch), так как вносит вклад в образование подъязычной кости. Остальные жаберные дуги обозначаются только номерами.
Мезенхима каждой бранхиальной дуги дает начало мышцам, хрящам и кости.
Клетки нервного гребня дают специфические скелетные структуры, например, эти клетки первой жаберной дуги дают Меккелев хрящ, верхнюю и нижнюю челюсть, различные связки и 2 из 3-х малых косточек, оссиклей, среднего уха. Типичная бранхиальная дуга содержит артерию (aortic arch), полоску хряща, мышечный компонент и нерв. (Рис.5.Б, Г-Е).
Кровь, проходящая через аортальные дуги вступает в дорзальную аорту (Рис.6.Д,Г).
Рис. 8. Рис. 8. А - область головы и шеи эмбриона человека (28 дней). Показаны зачатки мышц бранхиальных дуг. Стрелка показыват путь миграции миобластов из окципитальных миотомов для образования мускулатуры языка. Б - голова и шея плода человека ( 20 недель). Показаны мышцы, происходящие из бранхиальных дуг ( часть мышц platisma и sternocleidomastoid удалена, чтобы показать более глубокие мышцы). Миобласты из 2-й бранхиальной дуги мигрируют в область шеи и дают мышцы лица
1 - затылочные миотомы, 2 - m. orbicularis, 3 - m. buccinator, 4 - m. orbitalis oculi, 5 - m. frontalis, 6 - m. temporalis, 7 - v. occipitalis, 8 - m. auricularis, 9 - m. stylohyoid, 10 - m. stylopharingeus, 11 - anterior and posterior bellies of digastric, 12 - m. platisma, 13 - ключица, 14 - m. sternocleidomastoid, 15 - pharingeal muscles, 16 - m. mylohyoid, 17 - m. masseter
Хрящи бранхиальных дуг. Хрящевой компонент 1-й жаберной дуги называется Меккелевым хрящем (Meckel's cartilage), 2-й дуги - хрящем Рейхерта (Reichert's cartilage). Хрящи 3-й и 4-й дуг не имеют специального названия (см.29-й день, Рис.6). На рис.2 показано положение мышечных компонентов жаберных дуг (Рис.7.А) и их производных на голове и шее 20-недельного плода (Рис.7.Б).
А - вид сбоку на область головы и шеи у эмбриона человека 4-х недель. Показаны черепно-лицевые нервы, производные соответствующих бранхиальных дуг. Б - голова плода (20 недель). Показано поверхностное распределение двух каудальных ветвей тройничного нерва (V), производных первой бранхиальной дуги. В - сагитальный разрез через голову плода. Показано распределение глубоких сенсорных волокон бранхъиальных нервов к зубам, слизистой языка, глотке, полости носа, нёбу и гортани
1 - язык, 2 - нёбо, 3 - полость носа, 4 - паратидная железа, 5 - глотка, 6 - пищевод, 7 - гортань, 8 - низнечелюстная железа, 9 - подъязычные железы.
Состав внеклеточного матрикса
Компонент Хорда Аксиальная
мезенхима
Kлетки Оболочка 0 - отсутствие, (+) - слабая, + - сильная, ++ - очень сильная реакция
* - экспрессия, по-видимому, ограничена базальной мембраной Ходроитин/
дерматан сульфат 0 ++ +
Кератан сульфат 0 0 0
Гиалуроновая кислота 0 + +
Аггрекан 0 0 0
Бигликан + 0 0
Коллаген тип I (+), + (+) + <
Коллаген тип II 0 + 0
Декорин 0 ++ +
Фибронектин 0 ++ (+)
Ламинин (+) +* (+)
Тенасцин 0 + 0
TGF-β + > 0 0
Иммунореактивность и интенсивность окрашивания на 'TGF-beta увеличивается
Протеогликан Декорин обычно взаимодействует с коллагеновыми фибриллами, т,е.
участвует в фибриллогенезе, роль бигликана неизвестна. Коллаген тип II,по-видимому,
участвует в эпителиально-мезенхимных взаимодействиях (оболочка). Гликопротеин
фибронектин, по-видимому, связан с базальной мембраной и может выполнять
индуктивную роль. Молекулы ламинина, по-видимому, локализуются в
эндоплазматическом ретикулеме или аппарате Гольджи. Появление тенасцина в
оболочке хорды, по-видимому, связано с хондрогенезом зачатков тел позвонков и
межпозвоночных дисков, испытывающих преходящую хрящевую дифференцировку.
Секреция почти всех компонентов ВКМ в хорде человека регулируется TGF-beta. Так
TGF-beta индуцирует синтез декорина, который может связываться с фактором роста
через его core белок, модулируя его активность. Сходный механизм негативной петли
обратной связи обнаружен для ламинина.
В аксиальной мезенхиме межклеточные пространства заполнены
хондроитин/дерматан сульфатом и гиалуроновой кислотой. Они, особенно последняя,
ответственны за формирование гидратированных пространств, способбствующих
экспансии ВКМ ,миграции клеток и ингибированию дифференцировки или
хондрогенеза. След., склеротомные клетки иммигрирующие в аксиальную область
находят здесь идеальные условия для миграции и нахождения финальной позиции.
Этому способствует и фибронектин ВКМ аксиальной мезенхимы(Gotz et al., 1995).
Эмбрионы кур (Ст.) Эмбрионы мыши (Ст.)По Theiler's (1989) 16 стадия развития ,10-й день
В кровяных островках,образующихся на ранней сомитной стадии (Е8.0) ангиобласты все еще обнаруживают высокий уровень экспрессии flt-1, тогда как большинство гематопоэтических клеток экспрессирует flt-1
на низком уровне. Далее экспрессия flt-1 в гематопоэтических клетках снижается и наконец, экспрессия ограничивается только эндотелиальными клетками , достигая максимума к среднесомитной стадии (Е8.5). Это указывает на то, что гематопоэтические и эндотелиальные клетки происходят из общего предшественника. Затем экспрессия flt-1 в эндотелиальных клетках снижается и поддерживается на очень низком уровне на поздних стадиях эмбриогенеза.
Жаберный аппарат Развитие жаберных дуг начинается с миграции клеток нервного гребня (neural crest cells) в область будущей головы и шеи. В результате проникновения этих клеток образуется незначительное возвышение по бокам развивающейся глотки, первая бранхиальная дуга, зачаток челюстей. На 24-й день развития обнаруживается появление второй жаберной дуги , на 26-й день - третьей и на 28-й день - четвертой жаберной дуги. Ч-я и 6-я жаберные дуги малы и не могут быть видны на поверхности эмбриона. Жаберные дуги отделяются друг от друга хорошо заметными щелями, называемыми жаберными щелями (branchial grooves) (Рис.5А)
Рис. 7. Эмбрион человека (28 дней). Бранхиальный аппарат. Штриховыми линиями обозначены уровни разрезов
1 - , 2 - , 3 - , 4 - 1-4-я бранхиальные дуги, 5 - зачаток языка, 6 - Меккелев хрящ, 7 - первая дуга аорты, 8 - нерв, 9 - слепое отверстие, 10 - пищевод, 11 - ларинготрахеальная борозда, 12 - положение орофарингеальной мембраны, 13 - передний мозг, 14 - верхнечелюстное выпячивание, 15 - хорда, 16 - 1-й глоточный карман, 17 - сердце, 18 - рот, 19 - 3-я дуга аорты, 20 - дорсальная аорта, 21 - зачаток легкого, 22 - truncus arteriosus, 23 - 1-я бранхиальная мембрана, 24, 25 - 2-й и 3-й глоточные карманы, 26 - хрящ Рейхарта, 27 - мышцы, 28 - тироидный дивертикул
Первая жаберная дуга часто называется нижнечелюстной дугой (mandibular arch), она состоит из двух выпячиваний ("отростков") : (1) большого нижнечелюстного выпячивания (mandibular prominence), формирующего нижнюю челюсть, и (2) небольшого верхнечелюстного выпячивания ( maxillary prominence), формирующего верхнюю челюсть, скуловую кость (zygomatic bone) и чешуйчатую часть височной кости (tempotal bone) (Рис.7.В).
Вторая жаберная дуга часто называется гиоидной (hyoid arch), так как вносит вклад в образование подъязычной кости. Остальные жаберные дуги обозначаются только номерами.
Мезенхима каждой бранхиальной дуги дает начало мышцам, хрящам и кости.
Клетки нервного гребня дают специфические скелетные структуры, например, эти клетки первой жаберной дуги дают Меккелев хрящ, верхнюю и нижнюю челюсть, различные связки и 2 из 3-х малых косточек, оссиклей, среднего уха. Типичная бранхиальная дуга содержит артерию (aortic arch), полоску хряща, мышечный компонент и нерв. (Рис.5.Б, Г-Е).
Кровь, проходящая через аортальные дуги вступает в дорзальную аорту (Рис.6.Д,Г).
Рис. 8. Рис. 8. А - область головы и шеи эмбриона человека (28 дней). Показаны зачатки мышц бранхиальных дуг. Стрелка показыват путь миграции миобластов из окципитальных миотомов для образования мускулатуры языка. Б - голова и шея плода человека ( 20 недель). Показаны мышцы, происходящие из бранхиальных дуг ( часть мышц platisma и sternocleidomastoid удалена, чтобы показать более глубокие мышцы). Миобласты из 2-й бранхиальной дуги мигрируют в область шеи и дают мышцы лица
1 - затылочные миотомы, 2 - m. orbicularis, 3 - m. buccinator, 4 - m. orbitalis oculi, 5 - m. frontalis, 6 - m. temporalis, 7 - v. occipitalis, 8 - m. auricularis, 9 - m. stylohyoid, 10 - m. stylopharingeus, 11 - anterior and posterior bellies of digastric, 12 - m. platisma, 13 - ключица, 14 - m. sternocleidomastoid, 15 - pharingeal muscles, 16 - m. mylohyoid, 17 - m. masseter
Хрящи бранхиальных дуг. Хрящевой компонент 1-й жаберной дуги называется Меккелевым хрящем (Meckel's cartilage), 2-й дуги - хрящем Рейхерта (Reichert's cartilage). Хрящи 3-й и 4-й дуг не имеют специального названия (см.29-й день, Рис.6). На рис.2 показано положение мышечных компонентов жаберных дуг (Рис.7.А) и их производных на голове и шее 20-недельного плода (Рис.7.Б).
А - вид сбоку на область головы и шеи у эмбриона человека 4-х недель. Показаны черепно-лицевые нервы, производные соответствующих бранхиальных дуг. Б - голова плода (20 недель). Показано поверхностное распределение двух каудальных ветвей тройничного нерва (V), производных первой бранхиальной дуги. В - сагитальный разрез через голову плода. Показано распределение глубоких сенсорных волокон бранхъиальных нервов к зубам, слизистой языка, глотке, полости носа, нёбу и гортани
1 - язык, 2 - нёбо, 3 - полость носа, 4 - паратидная железа, 5 - глотка, 6 - пищевод, 7 - гортань, 8 - низнечелюстная железа, 9 - подъязычные железы.
Состав внеклеточного матрикса
Компонент Хорда Аксиальная
мезенхима
Kлетки Оболочка 0 - отсутствие, (+) - слабая, + - сильная, ++ - очень сильная реакция
* - экспрессия, по-видимому, ограничена базальной мембраной Ходроитин/
дерматан сульфат 0 ++ +
Кератан сульфат 0 0 0
Гиалуроновая кислота 0 + +
Аггрекан 0 0 0
Бигликан + 0 0
Коллаген тип I (+), + (+) + <
Коллаген тип II 0 + 0
Декорин 0 ++ +
Фибронектин 0 ++ (+)
Ламинин (+) +* (+)
Тенасцин 0 + 0
TGF-β + > 0 0
Иммунореактивность и интенсивность окрашивания на 'TGF-beta увеличивается
Протеогликан Декорин обычно взаимодействует с коллагеновыми фибриллами, т,е.
участвует в фибриллогенезе, роль бигликана неизвестна. Коллаген тип II,по-видимому,
участвует в эпителиально-мезенхимных взаимодействиях (оболочка). Гликопротеин
фибронектин, по-видимому, связан с базальной мембраной и может выполнять
индуктивную роль. Молекулы ламинина, по-видимому, локализуются в
эндоплазматическом ретикулеме или аппарате Гольджи. Появление тенасцина в
оболочке хорды, по-видимому, связано с хондрогенезом зачатков тел позвонков и
межпозвоночных дисков, испытывающих преходящую хрящевую дифференцировку.
Секреция почти всех компонентов ВКМ в хорде человека регулируется TGF-beta. Так
TGF-beta индуцирует синтез декорина, который может связываться с фактором роста
через его core белок, модулируя его активность. Сходный механизм негативной петли
обратной связи обнаружен для ламинина.
В аксиальной мезенхиме межклеточные пространства заполнены
хондроитин/дерматан сульфатом и гиалуроновой кислотой. Они, особенно последняя,
ответственны за формирование гидратированных пространств, способбствующих
экспансии ВКМ ,миграции клеток и ингибированию дифференцировки или
хондрогенеза. След., склеротомные клетки иммигрирующие в аксиальную область
находят здесь идеальные условия для миграции и нахождения финальной позиции.
Этому способствует и фибронектин ВКМ аксиальной мезенхимы(Gotz et al., 1995).
Эмбрионы кур (Ст.) Эмбрионы мыши (Ст.)По Theiler's (1989) 16 стадия развития ,10-й день
В кровяных островках,образующихся на ранней сомитной стадии (Е8.0) ангиобласты все еще обнаруживают высокий уровень экспрессии flt-1, тогда как большинство гематопоэтических клеток экспрессирует flt-1
на низком уровне. Далее экспрессия flt-1 в гематопоэтических клетках снижается и наконец, экспрессия ограничивается только эндотелиальными клетками , достигая максимума к среднесомитной стадии (Е8.5). Это указывает на то, что гематопоэтические и эндотелиальные клетки происходят из общего предшественника. Затем экспрессия flt-1 в эндотелиальных клетках снижается и поддерживается на очень низком уровне на поздних стадиях эмбриогенеза.
1 - , 2 - , 3 - , 4 - 1-4-я бранхиальные дуги, 5 - зачаток языка, 6 - Меккелев хрящ, 7 - первая дуга аорты, 8 - нерв, 9 - слепое отверстие, 10 - пищевод, 11 - ларинготрахеальная борозда, 12 - положение орофарингеальной мембраны, 13 - передний мозг, 14 - верхнечелюстное выпячивание, 15 - хорда, 16 - 1-й глоточный карман, 17 - сердце, 18 - рот, 19 - 3-я дуга аорты, 20 - дорсальная аорта, 21 - зачаток легкого, 22 - truncus arteriosus, 23 - 1-я бранхиальная мембрана, 24, 25 - 2-й и 3-й глоточные карманы, 26 - хрящ Рейхарта, 27 - мышцы, 28 - тироидный дивертикул
1 - затылочные миотомы, 2 - m. orbicularis, 3 - m. buccinator, 4 - m. orbitalis oculi, 5 - m. frontalis, 6 - m. temporalis, 7 - v. occipitalis, 8 - m. auricularis, 9 - m. stylohyoid, 10 - m. stylopharingeus, 11 - anterior and posterior bellies of digastric, 12 - m. platisma, 13 - ключица, 14 - m. sternocleidomastoid, 15 - pharingeal muscles, 16 - m. mylohyoid, 17 - m. masseter
1 - язык, 2 - нёбо, 3 - полость носа, 4 - паратидная железа, 5 - глотка, 6 - пищевод, 7 - гортань, 8 - низнечелюстная железа, 9 - подъязычные железы.
| Компонент | Хорда | Аксиальная мезенхима | |
|---|---|---|---|
| Kлетки | Оболочка | ||
| 0 - отсутствие, (+) - слабая, + - сильная, ++ - очень сильная реакция * - экспрессия, по-видимому, ограничена базальной мембраной | |||
| Ходроитин/ дерматан сульфат | 0 | ++ | + |
| Кератан сульфат | 0 | 0 | 0 |
| Гиалуроновая кислота | 0 | + | + |
| Аггрекан | 0 | 0 | 0 |
| Бигликан | + | 0 | 0 |
| Коллаген тип I | (+), + | (+) | + < |
| Коллаген тип II | 0 | + | 0 |
| Декорин | 0 | ++ | + |
| Фибронектин | 0 | ++ | (+) |
| Ламинин | (+) | +* | (+) |
| Тенасцин | 0 | + | 0 |
| TGF-β | + > | 0 | 0 |
В аксиальной мезенхиме межклеточные пространства заполнены хондроитин/дерматан сульфатом и гиалуроновой кислотой. Они, особенно последняя, ответственны за формирование гидратированных пространств, способбствующих экспансии ВКМ ,миграции клеток и ингибированию дифференцировки или хондрогенеза. След., склеротомные клетки иммигрирующие в аксиальную область находят здесь идеальные условия для миграции и нахождения финальной позиции. Этому способствует и фибронектин ВКМ аксиальной мезенхимы(Gotz et al., 1995).