ТЕЛОМЕРАЗА |
|
|---|---|
| Telomerase: Dr Jekyll or Mr Hyde? John C Prescott, Elizabeth H Blackburn Current Opinion in Genetics & Development 1999, 9:368-373. |
Теломераза большой ферментативный комплекс. У дрожжей Saccharomyces cerevisiae присутствует большая РНК субъединица (TER) примерно в ~430 kDa (1.3 kb), и по крайней мере две TER молекулы(и возможно две 103 kDa TERT молекулы) в теломеразном комплексе[12] [13]. Кроме того теломераза, по-видимому, содержит многочисленные другие белковые субъединицы. Только TER и TERT необходимы для основной каталитической активности [13] [14] [15]. Основная in vitro теломеразная активность может быть реконструирована по in-vitro-синтезированным TER и TERT, как для человеческой, так и Tetrahymena теломераз [16] [17•] [18].
References TLC1: template RNA component of Saccharomyces cerevisiaetelomerase. Science1994,266:404–409. Functionally interacting telomerase RNAs in the yeast telomerase complex. Genes Dev1997,11:2790–2800. Telomerase in yeast. Science1995,269:396–400. Three Ever Shorter Telomere (EST) genes are dispensable for in vitro yeast telomerase activity. Proc Natl Acad Sci USA1997,94:11190–11195. Reconstitution of human telomerase with the template RNA component hTR and the catalytic protein subunit hTRT. Nat Genet1997,17:498–502. Reconstitution of human telomerase activity in vitro. Curr Biol1998,8:177–180. The reverse transcriptase component of the Tetrahymena telomerase ribonucleoprotein complex. Proc Natl Acad Sci USA1998,95:8485–8490. Human telomerase RNA and telomerase activity in immortal cell lines and tumor tissues. Cancer Res1996,56:645–650. Selective inhibition of telomerase activity during terminal differentiation of immortal cell lines. Cancer Res1996,56:3796–3802. Differential regulation of telomerase activity and telomerase RNA during multi-stage tumorigenesis. Nat Genet1996,12:200–204. hEST2, the putative human telomerase catalytic subunit gene, is upregulated in tumor cells and during immortalization. Cell1997,90:785–795. Expression of TERT in early premalignant lesions and a subset of cells in normal tissues. Nat Genet1998,19:182–186. Telomerase activity in human ovarian carcinoma. Proc Natl Acad Sci USA1994,91:2900–2904. Extension of lifespan by introduction of telomerase into normal human cells. Science1998,279:349–352. Myc activates telomerase. Genes Dev1998,12:1769–1774. Dissociation among in vitro telomerase activity, telomere maintenance, and cellular immortalization. Proc Natl Acad Sci USA1998,95:14723–14728. Reconstitution of telomerase activity in normal human cells leads to elongation of telomeres and extended replicative life span. Curr Biol1998,8:279–282. Both Rb/p1 6INK4a inactivation and telomerase activity are required to immortalize human epithelial cells. Nature1998,396:84–88. Telomerase extends the lifespan of virus-transformed human cells without net telomere lengthening. Proc Natl Acad Sci USA1999,96:3723–3728. Absence of cancer-associated changes in human fibroblasts immortalized with telomerase. Nat Genet1999,21:115–118. Telomerase expression in human somatic cells does not induce changes associated with a transformed phenotype. Nat Genet1999,21:111–114. A mutant with a defect in telomere elongation leads to senescence in yeast. Cell1989,57:633–643. Sequence mutants of Saccharomyces cerevisiae with a defect in telomere replication identify three additional EST genes. Genetics1996,144:1399–1412. Telomere shortening associated with chromosome instability is arrested in immortal cells which express telomerase activity. EMBO J1992,11:1921–1929. Stabilization of short telomeres and telomerase activity accompany immortalization of Epstein-Barr virus-transformed human B lymphocytes. J Virol1994,68:3410–3414. Restoration of telomeres in human papillomavirus-immortalized human anogenital epithelial cells. Mol Cell Biol1994,14:961–969. Telomere length dynamics and chromosomal instability in cells derived from telomerase null mice. J Cell Biol1999,144:589–601. Telomerase RNA mutations in Saccharomyces cerevisiae alter telomerase action and reveal nonprocessivity in vivo and in vitro. Genes Dev1997,11:528–540. Telomere shortening and tumor formation by mouse cells lacking telomerase RNA. Cell1997,91:25–34. Essential role of mouse telomerase in highly proliferative organs. Nature1998,392:569–574. Severe growth defect in mouse cells lacking the telomerase RNA component. Nat Genet1998,19:203–206. Telomere shortening in mTR-/- embryos is associated with failure to close the neural tube. EMBO J1999,18:1172–1181. Evidence for an alternative mechanism for maintaining telomere length in human tumors and tumor-derived cell lines. Nat Med1997,3:1271–1274. An alternative pathway for yeast telomere maintenance rescues est1-senescence. Cell1993,73:347–360. Comparison of screening methods in the detection of bladder cancer. J Urol1999,161:388–394. Telomerase activity in gynecological tumors. Clin Cancer Res1996,2:2023–2028. Telomerase activity as a biomarker for (pre)neoplastic cervical disease in scrapings and frozen sections from patients with abnormal cervical smear. J Clin Oncol1998,16:2238–22345. Accelerated telomere shortening in young recipients of allogeneic bone-marrow transplants. Lancet1998,351:178–181. |
 Extension of cellular lifespan with telomerase expression Достаточность лишь TER и TERT для базальной теломеразной активности in vitro ставит вопрос, может ли эктопическая экспрессия теломеразной активности обеспечиваться эктопической экспрессией экзогенно добавленных генов для этих субъединиц. Хотя TER экспрессируется в большинстве типов тканей ее уровень экспрессии не коррелирует с уровнем экстрагируемой ферментативной активности [19] [20] [21]. С другой стороны, уровень экспрессии hTERT в некотрых тканях человека часто коррелирует с уровнем измеренной in-vitro теломеразной активностью и пролиферативным потенциалом ткани [22] [23] [24], указывая тем самым, что hTERT ограничивает in vivo формирование активного теломеразного комплекса. Показано, что конституитивной эктопической экспрессии hTERT достаточно для индукцмм теломеразной активности в клетках, в которых она не обнаруживалась [25••] [26] [27••] [28••] [29••] [30••].
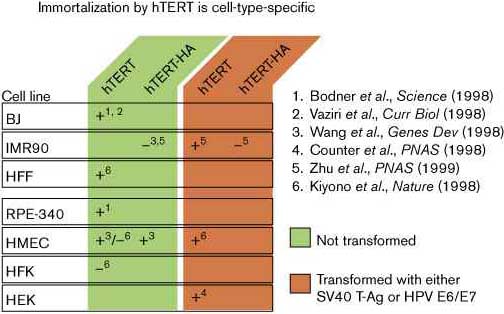
Эктопическая экспрессия hTERT в первичных культурах как в pre-senescent эпителиальных клетках (RPE-340) так и в pre-senescent фибробластах(BJ foreskin) была достаточна для индукции теломеразной актинвости и позволяла расти клеткам и за пределом нормальной репликативной способности, характерным для этих клеток (Fig. 2) [25••] [28••]. Более того, эти иммортализованные линии клеток не обнаруживали признаков хромосомных аномалий, типичных для post-senescent survivors [31••] [32••]. Вскоре было установлено, однако, что hTERT не достаточно для увеличения продолжительности жизни всех нормальных типов клеток. В одном исследовании [26] TERT экспрессия оказалась неспособной удлиннять жизнь легочных фибробластов человека (IMR90) — отметим, однако, что использованная hTERT конструкция содержала С-терминальный HA эпитопный тэг (epitope tag), см. ниже — однако продолжительность жизни клеток увеличивалась при экспрессии c-Myc который индуцировал эндогенную (преимущественно дикого типа) теломеразную активность. Было предположено, что необходимо второе трансформирующее событие (c-Myc expression) для теломеразы, чтобы увеличить продолжительность жизни этих клеток[26]. Эктопической экспрессии hTERT оказалось недостаточно и для увеличение периода жизни нетрансформированных молочных эпителиальных клеток человека (HMECs), содержащих, по-видимому, нормальный pRb/p16 путь ( pre-M1 [M0], HMECs), она оказалась неспособной иммортализовать и нормальные кератиноциты [29••]. Однако, ко-экспрессии вирусного HPV E7 белка, который инактивирует pRb, вместе с hTERT было достаточно для иммортализации как HMEC клеток, так и кератиноцитов. Факт, что большинство комерчески доступных HMECs являются post M0 может объяснить, почему в предыдущих исследованиях [26], один hTERT иммортализовал эти эпителиальные клетки [29••].
Telomerase activity in vitro versus telomere maintenance in vivo У дрожжей TER и TERT недостаточны для поддержания теломер или способности клеток к репликации in vivo. EST1, EST3, и EST4/CDC13 генные продукты дрожжей, хотя и не нужны для ферментативной активности теломеразы in vitro, являются существенными для поддержания теломер in vivo [14] [15] [33] [34]. В клетках человека конструкция hTERT , содержащая СМ-терминальный HA epitope tag поддерживает in vitro ферментативную активность и иммортализует HMEC клетки (which may have been post M0), однако неспособна поддерживать теломеры или преодолевать кризис в человечьих легочных (IMR90) и foreskin фибробластах, и в эмбриональных клетках человека (Fig. 2) [26] [27••] [30••]. Итак, в клетках дрожжей и человека каталитически активная теломераза недостаточна для поддержания теломер in vivo. Остается возможность, что HA tag в hTERT субъединице вовлечен во взаимодействие между TERT и предполагаемым человечьим гомологом EST1, EST3, или EST4/CDC13.
Поддержание теломер in vivo нуждается в продуктивном взаимодействии теломеразы с теломерным ДНК–белковым комплексом.
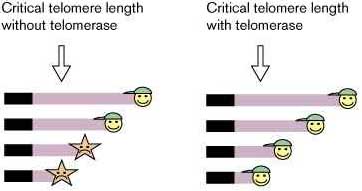
Telomere length and cell division capacity – how short is too short? Когда нормальные клетки растущие в культуре перестают делиться они в общем все еще содержат достаточное количество теломерной ДНК, следовательно, укорочение теломеры per se недостаточно для вызывания старения клетки. Клетки человека в культуре с репликативным периодом жизни увеличенном с помощью онкогеной трансформации продолжают укорочение теломер до тех пор пока не достигнут кризиса (Fig. 1) [46] [47] [48]. Нокаут гена TER мыши ведет к слиянию хромосом конец-в-конец, указывающему на compromised теломерную функцию, эти соединения часто содержат значительные количества теломерных повторов [49]. Тест способности теломеразы удлиннять продолжительность жизни post M1, SV40 T-Ag-трансформированных клеток, пролил новый свет на кажущийся парадокс. Эктопическая экспрессия hTERT в T-Ag-трансформированных HEK клетках удлинняет жизнь за пределы кризиса с соответствующим заметным удлиннением теломер [27••] [30••]. Экспресия hTERT (но не каталитически неактивного или carboxy-terminally HA-tagged hTERT) в T-Ag трансформированных фибробласах легких и кожи человека также удлинняет период жизни, несмотря на значительное укорочение теломер, которое продолжается десятки клеточных делений за пределами ожидаемого кризиса [30••]. Сравнительно высокий уровень теломеразной активности индуцирован в обоих исследованиях. Таким образом, удлиннение теломер per se необязательно для теломеразой-опосредованного удлиннения продолжительности жизни. Параллельно у S. cerevisiae, присутствие теломеразы с мутантной, хотя и функциональной, TLC1 РНК субъединицей стабилизирует теломеры на значительно более коротком уровне, чем у старых клеток, экспрессирующих стабильный, но энзиматически неактивный, теломеразный комплекс [50]. Таким образом, присутствие энзиматически активной теломеразы стабилизирует теломеры, которые могут стать критически короткими в ее отсутствие (Fig. 3).
Quality of life without telomerase Продолжительность жизни некоторых типов клеток может быть удлиннена с помощью экспресии hTERT, нокаутные мыши с отсутствием существенной РНК субъединицы теломеразы жизнеспособны и фертильны в течение более 6 поколонеий [51]. Последовательные возвратные скрещивания этих теломераза-дефицитных мышей выявляют постоянное снижение фертильности [52]. В то же самое время в соматических клетках увеличивается митотическая нестабильность, что видно по увеличению анеуплоидии и слияниям конец-в-конец хромосом [49] [52]. Эмбриональные стволовые клетки мыши с отсутствием mTER и поддерживаемые в культуре обнаруживают повышенные уровни слияний теломер и снижение репликативного периода жизни в культуре [53]. Наконец, поздние генерации mTER нокаутных мышей часто погибают in utero, у них часто отсутствует закрытие нервной трубки [54•]. Эти нарушения коррелируют с более короткой длиной теломер, чем у нормально развивающихся сиблингов ([54•]; M Blasco, personal communication).
TER нокаутные мыши, по крайней мере вначале, не более резистентны к определенным типам опухолей, чем мыши дикого типа [51]. Альтернативный (телдомераза-независимый) путь поддержания теломер, названный ALT, обнаружен в клетках человека. ALT ,по-видимому, активен примерно в ~10% опухолей человека с отсутствием обнаружимой теломеразной активности [55]. Этот второй путь поддержания теломер, по-видимому, сходен с RAD52-зависимым путем, обнаруженным ранее у почкующихся дрожжей — путем, который поддерживает теломеры с помощью неравной гомологичной рекомбинации [38] [56].
Однако большинство опухолей человека содержит повышенный уровень теломеразы. Это используется в клиниках для ранней диагностики опухолей. Анализ мочи для диагностики опухолей мочевого пузыря [57•]. Однако активность теломеразы может и не служить индикатором ранних стадий рака шейки [58] [59].
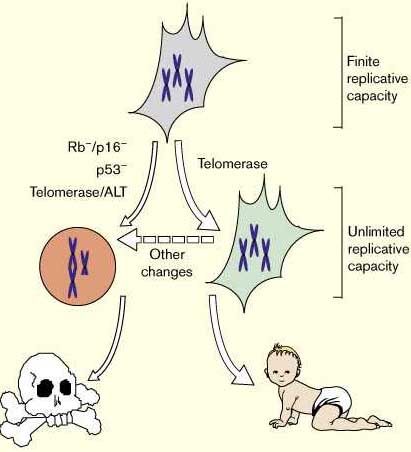
What price immortality? Недавние исследования показали, что теломераза способна иммортализовать некоторые типы клеток Fig. 4, такая иммортализация не сопровождается онкогенной трансформацией[31••] [32••]. Более того , геномная нестабильность является общей для канцерогенных клеток, так усиленная экспрессия теломеразы SV40-трансформированными фибробластами человека снижает появление дицентрических хромосом и гиперплоидии [30••], и таким образом защищает против, по крайней мере, одной причины геномной нестабильности. Теломеразой-обусловленная иммортализация клеток может таким образом служить терапевтическим средством в культуре растущих тканей человека.
Однако обратная сторона медали то, что эктопическая экспрессия теломеразы позволяет пре-неопластическим, частично трансформированным фибробластам, HMEC, и клеткам эмбриональных почек человека пролиферировать [27••] [30••]. В этом контексте экспрессия теломеразы обладает канцер-способствующими свойствами. Подсчитано, что после трансплантации костного мозга укорочение теломер может вызывать у донорских клеток старение на 15 лет [60]. С одной стороны такое "старение" устраняется эктопической экспрессией теломеразы (Dr Jekyll) в трансплантируемой ткани? С другой стороны, не будет ли экспрессия теломеразы (Mr Hyde) затем увеличивать вероятноть того, что трансплантированная ткань сама станет чувствительной к онкогенной трансформации ? ConclusionsНедавние исследования выявили Jekyll и Hyde природу теломеразы. Данные сумированные здесь показывают, что эктопическая экспрессия теломеразы может иммортализовать клетки без индукции геномной нестабильности, характерной для опухолевых состояний, и что эффекты эктопической экспрессии теломеразы являются клеточно-специфическими. Однако мы должны помнить о словах Robert Louis Stevenson's Dr Jekyll, ''I hesitated long before I put this theory to the test of practice. I knew well that I risked death; for any drug that so potently controlled and shook the very fortress of identity, might, by the least scruple of an overdose ... utterly blot out that immaterial tabernacle which I looked to it to change. But the temptation of a discovery so singular and profound at last overcame the suggestions of alarm'' [61]. |