В начале восьмидесятых годов, понимание физиологической функции улитки натолкнулось на противоречие: с одной стороны, нейрофизиологи описывали значительную избирательность в частотах на уровне волокон слухового нерва, с другой, теория волны, предложенная Bekesy
мешала рассматривать улитку, иначе чем в виде пассивного фильтра, неспособной отвечать за эту избирательность. Однако, в 1945 заявил об " активных механических процессах "! Два типа результатов помогли расшатывать тогда теорию "пассивности" улитки: и точные измерения вибрации органа Corti (мембраны basilaire) в ответ на чистый звук, которые показали, что избирательность в частотах осуществляется раньше, чем послание достигает слухового нерва.
При частототах чистого звука (здесь 10 khz, у морской свинки), активный механизм усиливает (env. + 50 db) вибрацию мембраны basilaire и главным образом он вызывает резонанс этой вибрации в очень небольшой порции органа corti, то, что делает значительной tonotopie (или избирательность частот). Этот резонанс в частотах ( tuning ) сильно зависит от целостности OHCs и передается на идентичном уровне волоконам слухового нерва, которым он передается и через IHCs.
Современная концепция электроподвижности
Электромобильность, осуществляемая OHCs, обнаружена в . Потом, это свойство сокращения (контрактильности) OHCs было широко изучено. Речь идет о быстром механизме (соответствующему высоким частотам (по крайней мере до 20-30 khz), который не зависит ни от кальция, ни от АТФ (он не потребляет, таким образом, энергии). Сразу же, исследователи выявили, что эта подвижность, вызываемая электрическим током, происходит на уровне ОНС. Сегодня считается, что деполяризация этой мембраны (поступление k+ во время стимуляции OHCs) изменяет строение белкового трансмембранного "двигателя", который обеспечивает посредством связи с худосочным подмембранным цитоскелетом сокращение клетки. OHC contraction is the sum of contraction of motors located within the OHC lateral plasma
membrane (ref.
e19). Then, because of the arrangement of this protein on the membrane, and because of its connection with
a cytoskeletal spring, shortening of the OHC length is induced. In turn,
the OHC coupling with support cells
allows transfer of this energy into the basilar and tectorial membrane,
giving the organ of Corti its exquisite properties of sensitivity and
frequency selectivity.
The current concept is that K+-mediated OHC membrane depolarisation
changes the conformation of the motor protein PRESTIN (see animated
drawing), which is now clearly identified (refs.
e22, e 23)
Schematic representation
of the OHC electromotility. (флеш-анимацмя)
OHC depolarisation
extracts anions (most probably chloride) from their prestin binding
sites. This induces a shortening of the prestin molecule and a contraction
of the plasma membrane. When the OHC repolarizes anions bind to prestin
again, which elongates.
Scheme:
G. Rebillard, animated by S. Blatrix
Ремарка
Сокращение OHCs ( электромобильность ) происходит на основе активного механизма, но он лишь достаточное условие. Чтобы сила, созданная этим преобразованием, переключила (электоромеханику) и была бы активна на целой партитуре улитки, необходимо кроме того тонкое между OHCs c другими структурами органа Сorti. Это соединение, которое осуществлено на базе градиента, убывающего от основания к апексу улитки, объясняет, почему активные механизмы были преобладающими для высоких и средних частот и значительно менее важны для низких частот.
Помимо электроподвижности, изолированные OHCs обнаруживают и некоторые другие контрактильные свойства: упомянем сокращение, которое регулирует электроподвижность in vivo .
Eсли у вас internet соединение, эта страница, подводя итог свойствам OHC сможет быть дополненной полезной информацией других сайтов:
, , , voir
aussi une concernant la
cochlée.
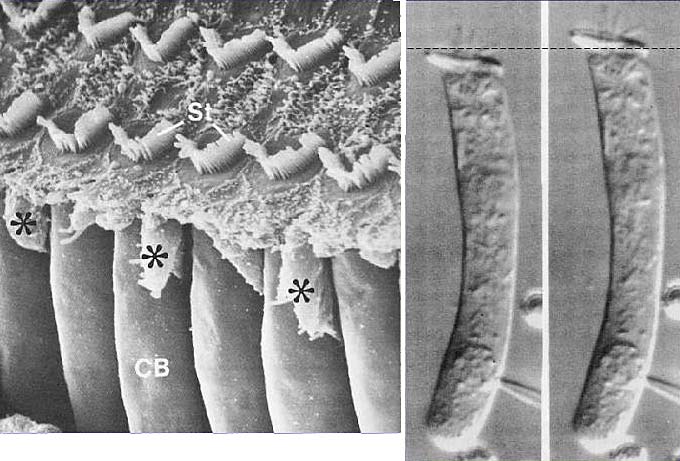
Наружные волосковые клетки (CB) находятся под эфферентным контролем головного мозга и помимо этого, они физически укорачиваются при электрической стимуляции (рис. справа). Изменение длины наружных волосковых клеток призвано усиливать чувствительность внутренних волосковых клеток путем увеличения вызываемой звуком shearing (давящей) силы, ощущаемой стереоцилиями внутренних волосковых
клеток от текториальной мембраны.
ELECTROMOTILITY
ORIGINAL RESEARCH PAPER Oliver, D. et al. Intracellular anions as the voltage sensor of prestin, the outer hair cell motor protein. Science 292, 2340-2343 (2001) | | |
LINK The authors' (requires QuickTime plug-in)
Идентифицирован - новый член семейства -related транспортеров, он обнаруживается в латеральной мембране OHC и является 'двигательным белком', отвечающим за OHC electromotility. Oliver и др. сообщили, что внутриклеточные анионы являются voltage сенсорами для prestin. Подробнее о престине ЗДЕСЬ
Когда звук проходит через улитку, вибрации преобразуются волосками на апикальной поверхности OHC, открывая механо-чувствительные ионный каналы. Возникающие в результате изменения мембранного потенциала индуцируют удлиннение клетки в случае гиперполяризации или укорочение, если клетка уже деполяризована. Эти motile реакции OHC сопровождаются prestin-зависимым движением зарядов, которые м.б. измерены экспреиментально.
Oliver и др. предположили, что заряженные аминокислоты в prestin скорее всего лежат в основе определяемого движения зарядов. Они вызывали мутации в наиболее вероятных кандидатах на роль незаряженных остатков и измеряли проходящий ток, однако характерные колоколо-образные электрические signature не были затронуты ни одной из мутаций.
Если voltage сенсор не является частью престина, то м.б. он является внешней (extrinsic) заряженной частицей. Они тестировали внутри- и внеклеточне ионы и нашли, что движение заряда фактически обусловлено движением цитозольных Cl- или HCO3- ионов через мембрану в ответ на изменение мембранного потенциала.
Согласно предложенной большинство молекул престина внедрено в латеральную мембрану(оболочку)OHC. Когда трансмембрнный потенциал меняется, то он индуцирует движение внутриклеточных анионов в или из кармана в престине, вызывая переключение между широкой и узкой конформацией. Если все молекулы престина находятся в узкой конформации, то клетки укорачиваются, если же они находятся в широкой , то клетки удлинняются. Престин является необычным 'motor protein', так как он не движется относительно точки прикрепления, а скорее меняет свой объем, и так как это происходит внутри плазматической мембраны, то они непрямо изменяют поверхность и, следовательно, длину клетки.