Sea Urchin Egg Endosomes Labeled with FM 1-43 and Tetramethylrhodamine-dextran.
После вступления в цитоплазму endocytotic пузырьки теряют клатриновую оболочку.
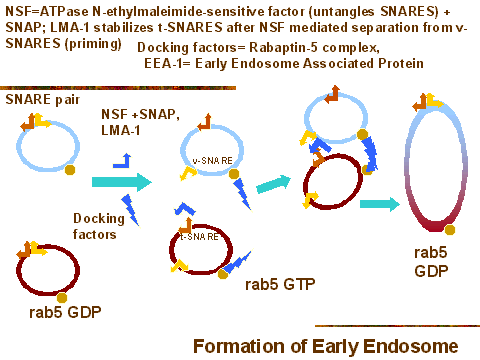
Они быстро сливаются с др. такими же пузырьками в процессе "гомотипического" слияния.
Каждый пузырек имеет пары SNARE белков, ( v-SNARE и t-SNARE) , которые используются
в инициальном процессе (и наз. cis-SNAREs). Гомотипическое слияние начинается с
разединения SNARE пар с помощью N-ethylmaleimide-sensitive factor (NSF) , который
использует также Soluble NSF attachment protein (SNAP).
Затем отсоединенные t-SNAREs стабилизируются с помощью др. фактора,
называемого LMA-1. Белок t-SNARE одного пузырька соединяется с v-SNARE др.
Тем временем, специфические docking/tethering факторы сводят пузырьки в
закрытый конгломерат, используя их поверхностные rab5-GDP. Один из них Rabaptin-5,
который появляется с nucleotide exchange factor (Rabex-5), который м. активировать rab5.
Кроме того имеются еще две GTPase , которые м. связывать две эндосомы вместе.
Еще один фактор - Early Endosome Associated Protein (EEA-1), который является
суперскрученными ( "coiled coils) спиралями. Его C конец имеет FYVE finger домен,
который связывает phosphatidylinositol-3-phosphate в эндосомных мембранах. EEA-1 не
только связывает эндосомы, он активирует слияние эндосом.
Т.обр., маркерами ранних эндосом являются:
EEA-1 белки
rab5-GDP
pH около 6.0
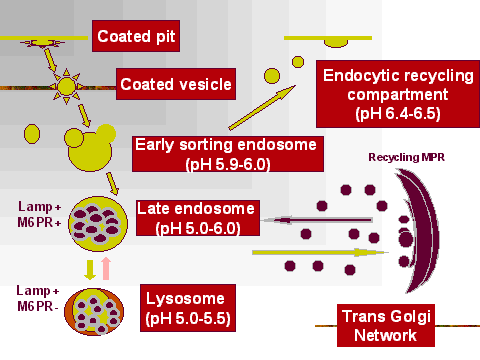
Ранние эндосомы имеют пониженный pH (5.9-6) и это позволяет освобождать
рецепторы и лиганды. Рецепторы м.б. использованы снова, они переносятся к поверхности с
помощью пузырьков, которые отшнуровываются от эндосом и затем направляются к плазматической
мембране. После этого recycling пузырьки сливаются с плазматической мембраной,
рецепторы возвращаются на клеточную поверхность. Затем ранние эндосомы превращаются
в поздние эндосомы.
Некоторые рецепторы м.б. деградированы. Некоторые переносятся в комплекс Гольджи,
чтобы вернуть их обратно в мембраны области Trans Golgi. Эти рецепторы подвергаются рециклингу. Этот процесс сходен с тем, с помощью которого происходит рециклинг lysosomal enzyme receptors. В большинстве случаев, как уже указывалось, рецепторы возвращаются обратно в плазматическую мембрану с помощью транспорта отпочковавшихся от эндосом пузырьков.
Formation of a late endosome
Поздние эндосомы образуются когда pH снижается до 5-6.0. Покрытые клатрином
пузырьки из Trans Golgi Network приносят переваривающие ферменты для поздних эндосом
и сливаясь с ними высвобождают свое содержимое. Поздние эндосомы становятся, таким образом,
degradative телами и приобретают кроме того маркер mannose 6 phosphate receptor "MPR+".
Они изменяют свой rab поверхностный маркер на rab7-GTP, возможно, давая пристанище
новымtargeting пузырькам, с которыми они сливаются. Т.обр., поздние эндосомы м. б.
идентифицированы по наличию rab7.
Поздние эндосомы включают мультивезикулярные тельца и содержат завитки или
пузырьки из мембран внутри. Они содержат также необычный липид, который становится др.
маркером для этой стадии. Липид называется lysobisphosphatidic acid (LBPA) , которая
имеет головнyю группу, большую, чем хвост. Ее структура способна проникать в сильно
искривленные мембраны, сходные с мембранами завитка. Считается, что это позволяет
удерживать и связывать специфические молекулы в завитках с помощью липид-протеиновых;
липид-липидных взаимодействий. Одним из таких типов молекул является холестерол, считается,
что это является важным местом для аккумуляции холестерола.
Поздние эндосомы функционируют, деградируя большинство белков и липидов.
Они также отвечают за возвращение MPR рецепторов обратно в сеть Trans Golgi.
Они рециклируют их путем отпочковываения мембран, которые переносят обратно рецепторы
для слияния их с мембранами Trans Golgi. После слияния, MRP рецепторы снова способны
отлавливать и сортировать новые деградационные энзимы для следующего переноса их в поздние
эндосомы.
Итак, поздние эндосомы содержат:
rab7-GDP
LBPA
MPR+
Late endosomes fuse with lysosomes
Наконец, поздние эндосомы м. оказаться неспособными переварить весь материал.
Тогда следующей ступенью является слияние их с лизосомами и создение гибридных органелл.
Оставшиеся сильно гликозилированные lysosomal associated membrane proteins (LAMPs) м. в
этом случает передаваться лизосомам. LAMPs затем становятся маркерами для поздних эндосом
или лизосом. Т.к. лизосомы не имеют MPR рецепторов (they have all been sent to the Golgi),
то это позволят отличать лизосомы и поздние эндосомы. Т.обр., слияние же начинается с того,
что MPR отсылаются назад в Trans Golgi.
Отметим, что лизосомы продолжают общаться с поздними эндосомами и м.
высвобождать важный материал обратно в эту групу органелл. Лизосомы рассматриваются как
конечный продукт эндоцитоза. Т.обр., лизосомы не общаются непосредственно с Trans Golgi
(а, следовательно, и с плазматической мембраной). Однако, они м. общаться с вышестоящими
структурами посредством ретроградного транспорта в поздние эндосомы.
Итак, маркерами
эндосом являются:
LAMP+
acid hydrolases
MPR negative
NPC1 (in normal cells)
THE ENDOCYTIC PATHWAY: A MOSAIC
OF DOMAINS
Jean Gruenberg Nature Reviews Molecular Cell
Biology 2, 721-730 (2001)
Органеллы в endocytic пути состоят из
мозаичных структур и функциональных
регионов. Эти регионы состоят, по
крайней мере частично, из
спеиализированных белково-липидных
доменов внутри плосткости мембраны
или белковых комплексов, ассоцированных
со специфическими мембранными липидами.
Тогда как некоторые из этих
молекулярных ансамблей обнаруживаются
в более чем одном компартменте, то
данная комбинация , по-видимому.
уникальна для каждого компартмента.
Следовательно организация мембраны м.б. modular.
В эндоцитотическом пути некоторые компартменты м.б. легко
идентифицированы благдаря их
мультивезикулярным характеристикам или
мультиламеллярному виду. Сходным
образом, Golgi полоска или сеть endoplasmic reticulum
м. обычно легко быть идентифицированы по
их типичной организации и топологии.
Границы между двумя легко различаемыми
компартментами оказываются размытыми
на молекулярном уровне, частично из-за
того, что ключевые белки, которые
регулируют мембранный транспорт часто
обнаруживаются в более, чем одном
компартменте. Разные мембранные
домены, которые м. обнаруживать
определенные биофизические свойства,
возможно, сосуществуют в каждом endocytic
компартменте. Динамическое
взаимодействие между этими доменами м.
создавать движущие силы, которые
отвественны как за специфическую
организацию каждого компартмента,
так и за перемещение груза молекул. Итак,
являются ли endocytic органеллы гомогенными
мембранными композициями?
Boxes
Box 1 | The endocytic pathways in yeast и mammals A degree of caution is needed when extrapolating from
mammalian to yeast endocytic pathways и vice versa, with
respect to the specific structure и organization of endosomal
membranes. One of the most striking features of mammalian
endocytosis, without discussing features of polarized или other
differentiated cells, is the capacity to reuse cell-surface
components efficiently и rapidly through recycling routes; whereas
equivalent recycling routes, if they exist, do not seem to be as
efficient in yeast cells. Although extensive sorting already occurs
in early endosomes of mammalian cells, it has been proposed that a
prevacuolar compartment equivalent to late endosomes functions as
the main sorting station in the yeast pathway.
In addition, yeast cells do not seem to share the striking
appearance of mammalian endosomes within the degradation pathway,
with a complex network of membrane invaginations. In yeast cells,
small и regularly shaped vesicles can be observed in the vacuole
of strains with impaired vacuolar hydrolase activity.
In addition to the basic machinery of yeast cells, mammalian cells
are likely to have evolved a more elaborate endosomal membrane
system to ensure optimal regulation и reutilization of proteins и
lipids.
В клетках высших эукариот
интернализация белков и липидов обеспечивается -coated пузырьками и др.
менее охарактеризованными путями.
Обычно эндоцитозируемые молекулы, включая
recycling рецепторы с их лигандами и downregulated
рецепторы, высвобождаются в ранние
эндосомы, где происходит эффективная
сортировка (Рис. 1). После разъединения рецептора и
лиганда при средней кислотности pH в просвете,
recycling рецепторы быстро (t1/2 ~ 2.5 min)
отделяются от своего лиганда и транпортируются вдоль маршрута
рециклинга, тогда как лиганды следуют по
пути деградации вместе с подавляемыми (downregulated)
рецепторами.
| Outline of the endocytic pathway.
Ранние эндосомы являются
динамическими компартментами с высоуой
способностью к гомотипическому слиянию.
Но их элементы оладают высокой
сложностью и плейоморфной организацией,
которая состоит из цистерновых областей,
от которых, по-видимому, отхдят тонкие
трубочки ( ~60 nm
диаметром) и большие пузырьки (~300–400 nm диаметром) (Рис. 2). Пузырьки содержат инвагинации в мембрнах и являются, следовательно,
мультивезикулами — хотя еще неясно до
какой степени эти инвагинации
отделяются от лимитирующих мембран и
образуют свободные пузырьки в просвете.
Тубулярные элементы рециклирующих
эндосом и мультивезикулярные
элементы напоминают endosomal
carrier vesicles/multivesicular bodies (ECV/MVBs) пути
деградации (Рис. 3). Эти тубулярные и
мультивезикулярные области м., следовательно, рассматриваться как
переходный вид (trans face) органелл.
Возможно, что центральная цистерновая
область функционирует как входная или cis
область, воспринимающая приближающиеся
пузырьки из плазматической мембраны или
сети trans-Golgi network.
Нелегко представить как мембраны
ранних эндосом м. формировать столь
разные структуры. Избирательные
изменения м. происходить на изгибах и
организации бислоя, напр., во время
инвагинаций мембраны при биогенезе молодых
ECV/MVBs или тубул. Морфогенез на
эндоцитотическом пути м. вовлекать в
действие молекулярные машины и
сегрегацию белков и липидов в плоскости
мембраны.
Sorting in early endosomes
Recycled и downregulated рецепторы эффективно
рассортировываются др от др. в ранних
эндосомах. Но трудно идентифицировать сортинг сигналы в
цитоплазматических доменах белков-грузов.
Это кажется неожиданным, т.к. такие
сигналы найдены для большинства, если не
всех, других ступеней транспорта
биосинтетического и эндоцитотического
пути. Не идентифицировано recycling
мотива, это ведет к предположению, что рециклинг
на клеточной поверхности происходит по
умолчаню. recycling to the cell surface occurs by default. Это
мнение трудно согласовать с ситуацией в
эпителиальных клетках, в которых transcytosed и
recycling рецепторы проходят через общие recycling
эндосомы прежде, чем они транспортируются к противоположным
доменам мембрнаны. Направляющие в
лизосомы сигналы идентифицированы, но
не всегда ясно на какой ступени
транспортировки они оперируют. Сортинг
мотивы найдены в цитоплазматических
доменах P-selectin , interleukin-2 (IL-2) receptor β chain, в вирусном белке Nef во время подавления CD4 и в epidermal growth factor receptor (EGFR), но эти мотивы мало походят
др. на др. Установлено, что ubiquitylation
цитоплазматических доменов м вносить вклад в лизосамальный сортинг. Временная monoubiquitylation белка также, как
полагают, действует на клеточной поверхности как интернализующий
сигнал и вполне возможно, что сходный
механизм оперирует при сортировке и
др местах клетки.
Кажущееся отсутствие сигналов к
рециклингу и разнообразие сигналов к
деградации указывают на то. что
различные принципы сортировки м.
оперировать в ранних эндосомах. Липиды
не распределены случайно внути
эндосомных мембран и разные липидные
домены м. сосуществовать на каждой
ступени пути. Рассортировка белков и
липидов — напр., во время биогенеза
тубул и ECV/MVBs — м.б. связана с избирательным
включением в специализированные
белок-липидные условия.
Привлекательными кандидатами для
выполнения сортировки в ранних
эндосомах являются липидные плотики-платформы
. Хотя судьба этих платформ в эндоцитозе неясна, предполагается, что
платформы на клеточной поверхности м.
действавать как интернализующие
платформы, маршрут, по которому следуют
преимущественно downregulated IL-2 рецепторы на
их пути к эндосомам и затем к лизосомам.
Кроме того, SV40 вступают в клетки через
кавеолы и затем располагаются в , которые не содержат endocytosed трассеров. Существуют ли функциональные
взаимоотношения между кавеосомами и
эндосомами, неизвестно. Компоненты
платформ, такие как cholesterol и sphingomyelin,
многочисленные в recycling эндосомах, по
крайней мере в CHO и Madin–Darby canine kidney (MDCK)
клетках, указывая тем самым, что
липидные платформы м. вносить вклад в
эндосомную сортировку. На самом деле, белки, которые
преимущественно включаются в платформы
на клеточной поверхности, следуют тому
же самому маршруту, что и сами платформы
в CHO клетках. Имеют ли компоненты
платформ то же самое распределние во
всех типах клеток, неизвестно. Более
важно установить, действительно ли эти
компоненты собираются в платформы
внутри эндосом — эндоцитозируемые
аналоги липидов с предпочтением к
доменам упорядоченных мембран
направляются в поздние эндосомы/лизосомы
скорее, чем в recycling эндосомы, в тех же
самых CHO клетках.
Molecular architecture of early endosomes
В дополнение к видимым мозаичным
ранним эндомомным доменам, становится
очевидным, что ключевые компоненты,
которые регулируют организацию мембран
и транспорт белков распределены не
случайно на мембранах, предопределяя
функциональные домены (Рис. 4). Могут ли пузырьки, прибывшие от
плазматической мембраны или сети trans-Golgi,
захватываться и сливаться где-либо с
этими мебмранами, неизвестно, но ели
ранние эндосомы обладают cis–trans
полярностью, то места docking/fusion д.б.
распределены неслучайно.
| Lipid–protein microdomains и molecular machines.
Docking/fusion белков семейства идентифицировано
вдоль обоих путей рециклинка и
деградации. Хотя данный SNARE м.б. наиболее
многочисленным в определенном
компартменте, но SNAREs неизбежно
распределяются по нескольим
компартментам во время везикулярного
транспорта. Итак, docking/fusion специфичность
не м.б. определена по распределению эндосомных SNAREs.
Более успешным оказалось изучение
малых GTPases семейства Rab , которые
рассматриваются как маркеры органелл из-за
их ограниченного распределения. В
активном GTP-связанном состоянии, Rab5, который участвует в раннем
эндоцитическом транспорте, м.
взаимодействаовать с несколькими
эффекторами. такими как др. Rab белки.
Благодая этим взаимодействиям, Rab5, по-видимому,
строит специфические эффекторные
платформы на мембране, которые м.
интегироватть различные механизмы,
которые регулируют транспорт, включая
слияние мембран, отпочковывание мембран
и взимодействие с компонентами
цитоскелета. Rab5 м. также вносить
вклад в пространственную организацию
мест docking/fusionдля пузырьков, прибывающих
разынми путями, т.к. его эффектор EEA1 м. взаимодействовать с syntaxin 6 , который участвует в
транспорте от trans-Golgiк ранним
эндосомам, и с syntaxin-13, который необходим для
слияния и рециклинга эндосом. Однако,
подобно др. ключевым регуляторам, Rab5
обнаруживается не только в одиночном
компартменте, но и ассоциирует с
плазматической мембраной, где его
активность, вообще связана с
образованием clathrin-coated пузырьков, по-видимому,
регулируемым путем передачи сигналов EGFR.
Установлено, что Rab5 эффектор, rabaptin 5, также взаимодействует с Rab4
— который участвует в рециклинге — и
что Rab4 эффектор, RABIP4, содержит FYVE motif и локализуется в ранних
эндосомах, следовательно, эти Rab
белки функционально связаны и отражают
суцествование физической связи между
разными платформами.
Некоторые белки взаимодействуют
специфически с phosphatidylinositol-3-phosphate (PtdIns(3)P) через законсервированный FYVEмотив,
включая Rab5 эффекторы EEA1 и rabenosyn-5, которые необходимы для
слияния ранних эндосом. EEA1 — и вообще др. FYVE белки — являются фактически ограниченными ранними эндосомами. Присутствие как
phosphatidylinositol 3-kinase (PI3K), так и PtdIns(3)P-связывающих белков среди Rab5 эффекторов должно облегчать
формирование микродоменов и платформ.
которые содержат Rab5 и PtdIns(3)P на мембранах
ранних эндосом. Хотя PtdIns(3)P–FYVE
взаимодействия достаточны для
нахождения ранних эндосом,
распределение FYVE белков также завист и
от др. компонентов. FYVE белок, hepatocyte growth factor-regulated tyrosine kinase substrate
(Hrs) — гомолог белка Vps27 — обнаруживается в ранних
эндосомах, даже когда FYVE мотив дефектен.
Кроме того, дрожжевой FYVE белок, Fab1, который является PtdIns(3)P-5'-OH киназой , которая генерирует
PtdIns(3,5)P2, является существенным для
поддержания нормальной морфологии
вакуолей и, как полагают, регулирует cargo-selective sorting
в просвете вакуолей. В дополнение к его
функции в динамикеранних эндосом, PtdIns(3)P
м. также функционировать в боигенезе
мультивезикулярных эндосом,
подчеркивая тесную связь между функцией
ранних жндосом и морфогенезом.
Выяволено существование и др.
phosphoinositide-связываюещего домена, общего
некоторым белкам, называемого PX-
или PHOX-homology доменом. Он впервые
обнаружен у p40phox и
p47phox — двух субъединиц neutrophil oxidase.
Домен PX у p40phox связывается с PtdIns(3)P
избирательно и присутствие этого домена
необходимо для эндоцитической функции и/или
локализации t-SNARE VAM7 и сортировки nexin SNX3. PX белок, SNX1, iвзаимодействует с FYVE белком
Hrs, и участвует в деградации EGFR, тогда как
дрожжевой омлолог SNX1, Vps5, является компрнентом , который
необходим для рециклинга
endosome–trans-Golgi network. Кроме того, SNX3 и SNX15 участвуют в эндосомном
транспорте, а их избыточная экспрессия
нарушает морфологию эндосом.
Взаимоотношения между PtdIns(3)P и PX белками или FYVE
белками все еще неясны.
Др. белки, которые м. функционировать в
организации функциональных областей в
мембранах, это annexins. Эти белки, по-видимому,
имеют врожденную способность к само-организации
на поверхности мембраны в двумерно
упроядоченные массивы.
Предполагается, что аннексины
используются в различных связанных с
мембранами механизмах, м. формирроваить
платформы на мембнанах рразных
субклеточных компартментов, которые
м. специфически взаимодействоваить с
цитозольными компонентами, включая цитосклет.
локализуется в
плазматической мембране и на ранних
эндосомах и участвует в динамике ранних и/или
recycling эндосом. Annexin II связывается с эндосомами, но не
с плазматической мембраной,
используя нестандартный холестерол-зависимый —
но кальций-независимый — механизм. Annexin II
взаимодействует также с белками
кортикального актинового цитоселета и
он, по-видимому. неслучайно распределен
на мембранах ранних эндосом и
концентрируется в областях, от которых,
по-видмому. исходят актин-подобные
филаменты. Следовательно, возможно, что
богатые холестеролом области мембран
ранних эндосом, которые взаимодействуют
с annexin II, важны для общей организации и
динамики этого компартмента.
coat комплекс регулирует
ретроградный транспорт от Гольджи в endoplasmic reticulum,
но имеется несколько указаний на то, что endosomal COPI complex, без β- и γ- subunits, участвует
эндосомном транспорте — особенно в биогенезе ECV/MVB
и Nef-опосредованной CD4 downregulation.
COPI кроме того участвует в транспорте от
фагосомных мембран. Подобно
биоситетическому COPI, эндосомный COPI
взаимодействует с мембранами через
малыую GTPase, ADP-ribosylation factor 1 (ARF1) . Однако, рекрутирование ARF1 и COPI
в эндосомы, но не в биоситетические
мембраны, зависит от кислотности
просветного pH, возможно благодая pH
сенсору, что ведет к связыванию ARF6 и ARF exchange фактора, ARNO. Как и в случае с др.
покровными компонентами, включая , а также clathrin и и
некоторые из их партнеров ,
COPI м. также взаимодействовать с
мембранными липидами. Таки слабые
взаимодействия м. облегчать таргетинг и
ассоциацию мембран и концентрировать
локально покрывные компоненты.
Тесные взаимоотношения между функцией
ранних эндосом в транспорте и
морфогенезе очевидны в функциях COPI.
Инактивация COPI в c температуро-чувстительными дефектами в ε-COP ингибирует
транспорт в поздние эндосомы и
взаимодействует с рециклингом transferrin
рецепторов, но не с рециклингом основной
массы, и нарушет также ранние эндосомы.
Они затем становятся тубулярными или
кластерами цистерн без мультивезикулярных доменов —
фенотип, напоминающий класс E VPS
мутантов у дрожжей. Идентичные
нарушения вызываются нейтрализацией
эндосомного pH, котора ингибирует связываение ARF1 и COPI.
Tubules of the recycling endosome
Два основных циркуита рециклинга и
деградации разделены как топологически,
так и функционально для гарантии, что
белки, необходимые для повторного
использования на клеточной
поверхности, будут отделены от тех. что
предназначены для деградации (Рис. 1). Однако, границы компартментов вдоль путей рециклинга и деградации, по-видимому,
размыты на молекулярном уровнеЮ в
особенности, если следить за одиночным
компонентом. Очевидно взаимодействие
между ключевыми компонентами, которые
определяют функциональные границы каждого пути.
Покинув ранние эндосомы участвующие в
рециклинге молекулы обнаруживаются
в отдельных тубулярных структурах,
которые соответствуют recycling эндосомам.
Они не содержат лигандов и рецепторов,
которые предназначены для деградации, и
они менее кислы, чем ранние эндосомы и
обнаруживаются вблизи центриолей в
некоторых, но не во всех типах клеток.
Эти тубулы формируют динамическую сеть
и вместе обозначаются как recycling эндосомы.
Rab5 ранних эндосом и две малые GTPases
участвуюющие в рециклинге, Rab4 и
, обнаруживают отдельные, но
частично перекрывающееся распределение
in vivo, которое преимущественно
соответствует разным эффекторным
платформам. Однако, точная функция Rab4 и Rab11
неясна. Члены семейства SNARE обнаружены в recycling
эндосомах, где они функционируют на пути,
включая v-SNAREs cellubrevin
и /VAMP8 в апикальном пути
поляризованных клеток, и t-SNARE syntaxin 13,
который участвует в слиянии эндосом.
Функция эти молекул неясна.
Рециклинг м. происходить с помощью
быстрого и медленного маршруов. Они
соответствуют, по крайней мере двум
раздельным ступеням транспорта в
плазматическую мембрану, каждый с
отдельной молекулярногй кухней (machinery);
напр., от ранних эндосом или от recycling
эндосом (Рис. 1). Альтерантивно, быстрый и медленный
транспорт м. отражать существование
градиента молекул на их пути к клеточной
поверхности внутри ранних и
recycling эндосом. Хим. в-ва или экспрессия
мутантых белков м. разбалансировать
транспорт вдоль любого пути и м.
объяснить, почему кинетика рециклинга жестко
зависит от деполимеризации
микротрубочек, т.к. перицентриолярное
распределение рециклинг-эндосом
зависит от микротрубеочек.
Транспорт вдоль пути рециклинга
зависит от актинового цитоскелета и
нестандартных миозиновых моторов,
которые м. выполнять механическую роль в
биогенезе и динамике тубул.
Взаимодействие между транспортом,
ремоделированием актина и Rac-опосредованной
передачей сигналов м. зависеть от малой GTPase ARF6 и
ее возможных партнеров.
Clathrin-coated почки обнаруживаются на recycling
эндосомах, но обеспечивадт ли они
транспорт назад на клеточную поверхность
или в trans-Golgi сеть, остается еще
определить. Т.к. клатриновые оболочки
обычно обладают сортирующей
способностью, то их присутствие на
recycling эндосомах подтверждает мнение, что
сортировка белков происходит внутри recycling
эндосом. Идентифицирован RME-1 — новый член
законсервированного семейства Eps15-homology (EH)-доменоых
белков, который обнаруживает
зарактеристики endocytic accessory белка. RME-1
ассоциирует с рециклинг-эндосомами и м.
участвовать в выходе мембранных белков
из этого компартмента.
Transport in the degradation pathway
Хотя тубулы обеспечивают рециклинг
для клеточной поверхности, транспорт же
промежуточных компонентов вдоль пути
деградации — от ранних к поздним
эндосомам — осуществляется ( ~300–400 nm diameter) ECV/MVBs. В клетках млекопитающих, ECV/MVBs четко отличаются от
ранних и поздних эндосом. В частности,
ECV/MVBs не содержат специфичных для ранних
эндосом белков или recycling рецепторов, они
не содержат также основные липидные и
белковые составляющие мембран
поздних эндосом. Однажды сформированные, ECV/MVBs
движутся в направлении поздних эндосом
микротрубочки- и мотор-зависимым
способом, и обладают способностью
закрепляться на и сливаться
избирательно с поздними эндосомами,
ступень, которая зависит от стандартной docking/fusion
кухни (machinery). В аксонах, ECV/MVBs также
функционируют как передаточные
структуры от ранних эндосом в
пресинаптических мембрнах к поздним
эндосомам в теле клетки. Меняеется ли
композиция ECV/MVBs, когда они
подвергаются процессу созревания
или когда они обеспечивают транспорт
между двумя стабильными компартментами?
ECV/MVBs, подобно поздним эндосомам,
содержат два морфологически видимых
мембранных домена, внутренние
инвагинации и ограничивающие мембраны.
Состав последних неизвестен, за
исключением того, что они, по-видимому не
содержат белков ранних эндосом и
отличаются от мембран поздних эндосом,
которые содержат высокие количества LAMP1. Также мало известно и о
композиции ECV/MVB инвагинаций. Они накапливают downregulated
рецепторы, в частности EGFR, это позволяет
предположить. что внутренние
мембраны предназначены к деградации
с помощью лизосом. ЭМ исследования
показали. что PtdIns(3)P многочисленны во
внутренних мембранах ECV/MVB, помимо ранних
эндосом, и что они не обнаруживаются
внутри поздних эндосом, которые
содержат (LBPA). Однако, неясно, присутствие этих PtdIns(3)P на
внутренних мембранах отражает роль этих
липидов в биогенезе инвагинаций или
просто в метаболизме липидов.
Wortmannin, в-во, которое ингибирует PI(3)K ,
ингибирует и формирование
мультивезикулярных тел в клетках
млекопитающих. В то же время
накапливаются доказательства у дрожжей,
что передача сигналов через PI(3)K Vps34 и PtdIns(3)P является ключевой в
транспорте вакуолей из trans-Golgi
сети. Учитывая эти тесные
взаимоотношения между ECV/MVB биогенезом и
организацией ранних эндомом, м. ожидать ,
что некоторые компоненты, которые
регулируют функции ранних эндосом ,
также важные для процесса инвагинации.
Кроме того,мутации дрожжей класса E VPS
дефектны по биогенезу MVB, т.к. они
формируют кипы трубочек или
искривленные цистерны, вообще в
процессе 'frustrated invagination'. Некоторые
классы E мутантов bv.n гомологов у
млекопитающих. которые вовлечены в
транспорт, включая Vps27,
Vps23 и Vps4 гомологи, iуказывая тем
самым, что функция этих белков м.б.
законсервирована в клетках млекопитающих (Box
1). Предполагается. что дрожевой
компартмент класса E — или превакуолярный
компартментt — соответствуют поздним
эндосомам, но их аналоги у млекопитающих
остаются неизвестными (Box
1). клетках млекопитающих фенотип,
который морфологически сходен с
фенотипом класса E мутантов еще
охарактеризованы недостаточно, за
исключением ранних эндосом после COP
инактивации в LDLF клетках,
или Hrs разрушения у мышей.
The pomegranate и the onion
Ранние эндосомы и элементы поздних
эндосом оченть динамичны, с очень
сложной и плейоморфной организацией,
содержащей цистрновую, тубулярную и
везикулярную области с многочисленными
инвагинациями мембраны. Их
ограничивающие мембраны сходны с
таковыми лизосом, содержат высокие
количества LAMP1, которые, как полагают,
защищают от деградирующей среды
компартмента благодрая их вычтокой
степени гликозилирования. Лимитирующие
мембраны поздних эндосом содержат
также MLN64, гомологи steroidogenic
acute regulatory protein (StAR). Внутренние мембраны клеток
высших эукариот подвергаются значительному ремоделированию и
накапливают большие количества ( ~15% от всех phospholipids) LBPA, которые являются чистым субстратом для phospholipase и
следовательно. резистентны к деградации.
LBPA не обнаруживаются в наружной
поверхности лимитирующего бислоя. До
какой степени биогенез и динамика
внутренних мембран поздних эндосом
регулируются с помощью одних и тех же
механизмов, которые оперируют во время
отпочковывания ECV/MVBs, неясно. LBPA , по-видимому,
синтезируется in situ
и имеют форму перевернутого конуса. Эта
структура м. облегчать инвагинации,
которые образуются мультивезикулярными
элементами поздних эндосом.
Белки, которые предназначены для
деградации, накапливаются в ECV/MVB
внутренних мембранах. Это позволяет
предположить, что их судьба быть
деградированными в лизосомах. Было
показано, что негативно заряженные
фосфолипиды, в частности LBPA, значительно
облегчают деградацию некоторых
гликолипидов. Т.к. LBPA сам деградирует
слабо, то его единственной функцией в
мембрнах - представлять липиды и белки,
которые нужно деградировать с помощью
гидролитической machinery.
Однако, внутренние мембраны поздних
эндосом не только содержат белки,
который предназначены для деградации.
Члены , включая CD63/LAMP3, накапливаются внутри
внутренних мембран, помимо их присутствия на клеточной
поверхности. В , major
histocompatibility complex (MHC) class II compartments (MIIC)
обладают большинством характеристик
поздних эдосом и лизосом, включая
сложную систему внутренних мембран, правда они являются все же
специализированными компартментами,
содержащими с специфические компоненты,
такие как cathepsin S protease, необходимая для
процессинга антигенов
и human leukocyte antigen DM (HLA–DM) комплекс.
MHC class II молекулы многочисленны в MIIC
компартменте как в В лимфоцитах, так и в
незрелых . MHC class II молекулы ко-локализуются с LBPA в B лимфоцитах и
они обнаруживаются также в LBPA-содержащи[
микровезикулах , преимущественно
секретируемых antigen-presenting клетками.
Наконец, молекулы (M6PR) транспортируемые между
trans-Golgi network и поздними эндосомами для
высвобождения лизосомных энзимов в
эндосомах и лизосомах. При транзите
через поздние эндосомы M6PR
преимущественно обнаруживается во внутренних мембранах.
LBPA мембраны вероятно выполняют важную
функцию по транспорту через поздние
эндосомы. Так, эндоцитозированные in vivo,
антитела против LBPA накапливаются в
поздних эндосомах и специфически
ингибируют транспорт M6PR. Напротив,
потеря экспрессии M6PR способствует
накоплению LBPA в виде мультиламеллярных
тел. Зндоцитозированные анти-LBPA
антитела вызывают также накопление
холестерола в поздних эндосомах,
воспроизводя болезнь накопления
холестерола Niemann–Pick type C (NPC), это указывает
на то, что LBPA мембраны служат для сбора и
распределения устройств для low-density
lipoprotein (LDL)-derived холестерола. В свою
очередь накопление холестерола в
поздних эндосомах взаимодействует с
циклом M6PR в некоторых типах клеток, а также с
циклом CD63 и p-selectin в тела в
эндотелиальных клетках. Sphingolipids
накапливаются в мультивезикулярных
компартментах в тканях пациентов с NPCи,
напротив, холестерол накапливается в в
поздних эндоцитических компартментах
при некоторых б-нях накопления сфинголипидов.
Функция мемран LBPA в транспорте белков
и липидов указывает на то, что некоторые
мембранные белки и липиды не
проникают во внутренние мембраны, а м.
постепенно выходить и возвращаться в
другие клеточные места назначения.
Включение во внутренние мембраны м.б.
легко объяснено селективным
разделением на части некоторых молекул
внутри LBPA инвагинаций или или
селективным исключением из
лимитирующих мембран. Если это так, то
экспорт из поздних эндосом д.б.
медленным,т.к. внутренние мембраны
многочисленны и лишь молекулы,
присутствующие на ограничивающих
мембранах м.б. пригодными к транспорту (Рис. 5) — это м. б. объяснить медленный цикл CD63 в эндотелиальных клетках. Др. механизмы,
однако, м.б. призваны объяснить
эффективность экспорта — напр.,
транспорт M6PR или экспорт MHC class II в
плазматическую мембрану в активированных дендритных клетках.
Единственная возможность, что транспорт
между внутренней и ограничивающей
мембранами регулируется с помощью
мехинизмов, включая постртансляционные
модификации и, в ча тности,
ubiquitylation. Возможно также, что поздние
эндосомы в клетках высших эукариот м.
содержать более одного типа внутренних
мембран, таких как те, что
специализированы для деградации белков
или рециклинга. Это согласуется с
находками, что ECV/MVBs и поздние эндосомы
отличаются по своему LBPA содержимому,
хотя оба имеют многоцисленные
внутренние мембраны. Более того, поздние
эндоцитические компартменты в
клетках млекопитающих м.выглядеть
подобно гранату (multivesicular)
или подобно луковице (multilamellar). Ничего
неизвестно о функциональном значении
этих различных типов внутренних мембран
или о различиях их биоситетического
состояния, организации и состава. М.
предположить. что помимо основной кухни
( machinery), которая ответственна за
образование внутрипросветных пузырьков
у дрожжей — которая должна
зависеть от класса E VPS белков и PtdIns(3)P
сигнализации — клетки млекопитающих
используют более эффективные механизмы
рециклинга, вообще LBPA зависимые, для
белков, которые нуждаются в экспорте из
внтуренних мембран поздних
эжндоцитических компартментов.
| Degradation или recycling?
The end point или a dynamic network?
Трудно провести линию между ранними и
поздними эндосомами, еще труднее между
поздними эндосомами и лизосомами.
Последние оба содержат лизосомные
жнзимы, их pH одинаково кислое ( ~5.5), и их
ограничивающие мембраны в первую
очередь состоят из одних и тех же
гликопротеинов. Лизосомы м.б.
идентифицированы только по их
физическим параметрам в градиентах и по
их ЭМ виду и по томуФ. что они не
содержать белков, обнаруживаемых в
поздних эндосомах, включая временные M6PR, Rab7 и Rab9, или фосфорилированные
предшественники hydrolase. Итак, не выявлено
ни белков, ни липидов, которые
присутствовали только в лизосомах, но не
в поздних эндосомах, кроме тогоу них
общая и базовая docking/fusion
machinery. Более того, поздние эндосомы и
лизосомы м. обмениваться содержимым
и мембранными белками быстро и
эффективно и они, по-видимому.
динамически взаимодействуют образуя
гибридные проежуточные структуры.
Предполагается, что поздние эндосомы и
лизосомы соответствуют отдельным
элементам общей динамической сети.
такая суб-компартментализация на
регионы и мембраные домены, которые
отличаются структурно и функционально,
напоминает организацию ранних эндосом.
Высоко подвижные тубулярные лизосомы
макрофагов м представлять собой более
стабильную или крайнюю версию такой
сети, которая согласуется с тем фактом,
что они обладают некоторыми общими с
поздними эндосомами характеристиками.
LBPA мембраны м. обладать фунцией таможни
в этой сети, т.к. они, по-видимому,
вовлекаются в транспорт как белков, так
и липидов, а также в деградацию липидов.
М. существовать и др. специализированные
области. Это м.б. объяснить, что
различные популяции поздних эндосом
сортируют и упаковывают молекулы груза
спосощью Rab9–TIP47 или с помощью retromer комплекса,
а также присутствие специализированных
поздних эндосом или лизосом в высоко
дифференцированных клетках, включая MIIC
в antigen-presenting клетках.
Экспорт холестерола м. происходить в
специализированных элементах, т.к. NPC1 белок — который вовлекается
во внутриклеточный транспорт
холестерола — распределен в
субнаборе лизосом. Нарушения
транспорта NPC и др. б-ни накопления
липидов м.б. результатом коллапса
мозаичной архитектуры эндосом, ведущей
к перемешиванию липидно-протеиновых
доменов иингибированию трансопрта.
Conclusions
Итак, ранне эндосомы содержат Rab5 и его
эффекторы, PtdIns(3)P и партнеры, эндосомные COPs,
компоненты платформ (raft) и annexin II, все
они участвуют в транпорте через этот
компартмент. Каждая из этих молекул
обнаруживается также и в др. мембранах,
но эта специфическая комбинация имеется
только в ранних эндосомах. М.
предположить, что эндосомные мембраны и
очевидно мембраны др. органелл строятся
из модулярных элементов. Это м.
объяснить трудности выявляения границ
определенных органелл на молекулярном уровне.
Необходимо понять как эти разные
мембранные машины и динамические домены
интегрируют для поддержания транспорта
белков и липидов, а также структуры и
функции органелл. Установлено
существование тесного взаимодействия между путями
передачи сигналов и транспорта через
белковую сеть.
 ЭНДОСОМЫ
ЭНДОСОМЫ 
 | Outline of the endocytic pathway.
| Outline of the endocytic pathway.  | The early
endosome.
| The early
endosome.  | Endosomal carrier vesicles/multivesicular bodies.
| Endosomal carrier vesicles/multivesicular bodies.  | Lipid–protein microdomains и molecular machines.
| Lipid–protein microdomains и molecular machines.  | Degradation или recycling?
| Degradation или recycling?