Lundblad V, Wright WE:
Telomeres and telomerase: a simple picture becomes complex. Cell1996,87:369–375.
Nakamura TM, Cooper JP, Cech TR:
Two modes of survival of fission yeast without telomerase. Science1998,282:493–496.
Promisel Cooper J, Nimmo ER, Allshire RC, Cech TR:
Regulation of telomere length and function by a Myb-domain protein in fission yeast. Nature1997,385:744–747.
van Steensel B, de Lange T:
Control of telomere length by the human telomeric protein TRF1. Nature1997,385:740–743.
•
van Steensel B, Smogorzewska A, de Lange T:
TRF2 protects human telomeres from end-to-end fusions. Cell1998,92:401–413.
Krauskopf A, Blackburn EH:
Control of telomere growth by interactions of RAP1 with the most distal telomeric repeats. Nature1996,383:354–357.
Smith S, Giriat I, Schmitt A, de Lange T:
Tankyrase, a poly(ADP-ribose) polymerase at human telomeres. Science1998,282:1484–1487.
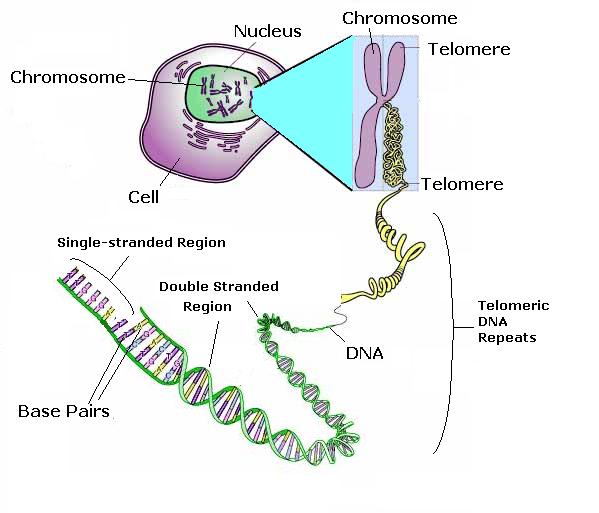
Теломеры не реплицируются полностью с помощью ДНК полимераз и постепенно укорачиваются с каждым клеточным делением [3]. Когда теломеры становятся короткими хромосмы становятся митотически нестабильными и ведут к случайным анеуплоидиям, а иногда и к гибели клеток[4][5].
Теломераза, рибонуклеопротеиновая обратная транкриптаза, которая синтезирует одну нить теломерной ДНК и противодействует тем самым потере теломерной ДНК [6].
Макромолекулярные компоненты теломеразы и ее ферментативная активность идентифицированы у разных организмов[7]. Активность теломеразы не обнаруживатеся в большинстве соматическийх тканей челоека, но она постоянно обнаруживается в высоко репликативной ткани и в большинстве опухолей человека. [8].
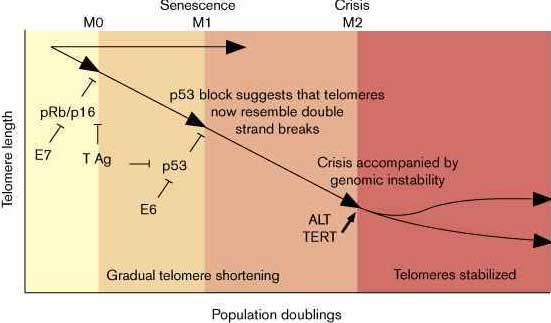
Известно, что у эукариот при разрушении теломеразы теломеры укорачиваются и клетки перестают делиться после нескольких десятков делений [9]. Hayflick наблюдал, что нармальные соматические клетки человека обладают ограниченной репликативной способностью (see [4][5][10] for review). Клетки могут преодолеть предел старения или M1, когда они трансформируются каким-либо из онкогенов, таким как SV40 T-Ag или HPV E6/E7. Такие трансформированные клетки продолжают делитьтся пока не достигнут второго блока пролиферации, названного crisis (M2). Теломеры прогрессивно укорачиваются в течение всего процесса репликативного клеточного старения ('aging'), предполагается, что теломеры действуют как митотические часы ('mitotic clock')[11]. Эта гипотеза подтвержадется наблюдением, что мноuие соматические ткани не содержат обнаружимых теломер, тогда как большинство иммортализованных или опухолевых клеток имеют их. Канцерогенные клетки, прошедшие М1 и избежавшие кризиса теряют контроль над клеточными циклами, имеют нестабильный геном и формируют метастазы in vivo, и обычно имеют довольно стабильные, хотя и разной длины теломеры(Fig. 1) [3].
Рис. 1 Когда клетки человека пролиферируют в культуре, то наступает пролиферативная блокада. Некоторые клетки находятся в pRb/p16-зависимом аресте клеточного цикла (M0), который останавливает пролиферацию после десяти или меньшего числа удвоений клеток. Независимо от M0 ареста клетки затем подвергаются p53-зависимому аресту (M1) после 40–50 делений. Клетки, которые проходят M1 подвергаются финальному блоку после дополнительных 30–40 делений(M2). Эти пролиферативные критические точки могут быть преодолены с помощью ряда вирусных онкогенов E7, T Ag, E6, которые инактивируют pRb/p16 и/или p53, и с помощью стабилизации теломер или теломеразой (TERT) или ALT.
Figure 1 As human cells proliferate in culture, telomere lengths shorten and the cells are subject to a succession of proliferative blockades. Some cells exhibit a pRb/p16-dependent cell cycle arrest (M0) that stops proliferation in culture after ten or fewer cell doublings. Whether cells exhibit an M0 arrest or not, they are subject to a later p53-dependent arrest (M1) after 40–50 cell doublings. Cells that bypass M1 are then subject to a final block to immortality following an additional 30–40 doublings (M2). These proliferative checkpoints can be bypassed by a number of viral oncogenes E7, T Ag, E6 that inactivate pRb/p16 and/or p53, and by telomere stabilization by either telomerase (TERT) or ALT.
Повреждение компонентов или теломеразы или самой теломеры меняет регуляцию длины теломер, что ведет к важной концепции, что гомеостаз длины теломер связан с регулирующим доступом теломераз к теломерам [35][36][37][38][39]. Присутствие теломераз в клетке не гаранитирует ее доступ к теломере.
Гомеостаз теломер регулируется, по крайней мере частично, теломер-связывающими белками: обильными Rap1p в почкующихся дрожжах, Taz1p в делящихся дрожжах и TRF1 и TRF2 у человека, все они специфически связываются с повторами дуплексной теломерной ДНК [40][41][42][43•]. Теломеры удлинняются в ответ на увеличение числа Rap1p молекул и других структурных признаков комплекса, присутствующего на конце хромосомы ([35][36][37][44]; CD Smith, J Prescott, EH Blackburn, unpublished data). Кроме того, мутантные теломерные повторяющиеся последовательности редуцируют сродство Rap1p к теломере и могут дерегулировать процесс поддержания длины теломер ([35][37][44]; CD Smith, J Prescott, EH Blackburn, unpublished data). Такие мутации не нарушают уровень теломеразной активности in vitro (T Boswell-Fulton, J Prescott, EH Blackburn, unpublished data). Сходным образом, мутации TRF1 , которые нарушают его связывание с теломерой, ведут к удлиннению теломер [42]. Присутствие TRF1 на теломерной ДНК может регулироваться в клетках с помощью Tankyrase (TRF1-interacting ankyrin-related ADP ribose polymerase), недавно идентифицированного энзима, который добавляет poly(ADP–ribose) как к себе, так и к TRF1 in vitro,снижая тем самым сродство TRF1 к теломерной ДНК [45]. Такая модификация может служить сигналом для удлиннения теломер.
ORIGINAL RESEARCH PAPER
Bailey, S. M. et al. Strand-specific postreplicative processing of mammalian. Science 293, 2462-2465 (2001) | Article | PubMed | ISI |
Линейные хромосомы несут уютно пристроившиеся нуклеопротеиновые шапочки (cap), которые предупреждают деградацию хромосомных концов и защищают от несоответствующей рекомбинаии. В клетках млекопитающих эти шапочки - теломеры - представлены повторяющимися G-rich последовательностями, связанными с рядом белков, включая Ku70, Ku80, каталитическую субъединицу DNA-dependent protein kinase (DNA-PKcs) и telomeric-repeat binding factor 2 (TRF2). Goodwin и др. обсуждают, как процессинг теломер связан с их способом репликации.
Two TCCs in HTC75 cells. The tel-G probe (which detects leading-strand telomeres) was hybridized and photographed, then the probe removed by denaturation and the tel-C probe hybridized and photographed. Yellow arrowheads indicate the point of fusion. Image courtesy of Michael Cornforth, University of Texas. (Cen, centromere.)
Для изучения механизма capping авт. использовали негативные мутанты TRF2 (TRF2BM), у которых удалены эндогенные TRF2 из теломер. Они экспрессировали TRF2BM в течение 5 дней в HTC75 клетках fibrosarcoma человека и нашли, что 44 из 154 митотических клеток обнаруживают конец-в-конец слияния хромосом. Эти слияния - подогнанные telomeric chromatid concatenates (TCCs) - образуются за счет участия одной из сестринских хроматид от каждей сливающейся хромосомы (Рис.), указывая тем самым, что TRF2 capping, м. происходить после репликации теломер.
Шапочка должна не только отделитьтся, чтобы произошла репликация, но и должна также повторно сформироваться после этого. Репликация приводит к генерации двух новых теломер - одна продуцируется с помощью синтеза leading-нити ДНК, др. с помощью синтеза lagging-нити.
Goodwin и др., используя хромосом ориентированную флюоресцентную in situ гибридизацию, которая дает различные гибиридизационные паттерны в зависисмости от типа слияния - слияния lagging-lagging, lagging-leading или leading-leading нитей. Они нашли что у TRF2BM в 133 из 135 случаев, TCCs продуцировалось в результате слияний между leading-strand теломер.
Почему? Goodwin и др. полагают, что это м.б. обусловлено тем, что концы генерируются с помощью двух способов репликации - leading нити заканчиваются тупо, т.к. имеют абсолютную потребность в TRF2 и DNA-PKcs чтобы fashion 3' выступал до образования t-петли на хромосомном конце. Lagging-strand теломеры, с др. стороны, уже имеют 3' выступ после репликации. Возможны и др отличия в способе образования двух типов теломер.
ORIGINAL RESEARCH PAPER Baumann, P. & Cech, T. R. Pot1, the putative telomere end-binding protein in fission yeast and humans. Science 292, 1171-1175 (2001) | Article | |
FURTHER READING McEachern, M. J. et al. Telomeres and their control. Annu. Rev. Genet. 34, 331-358 (2000) | |
Теломеры - белок-ДНК комплексы на концах хромосом - защитные устройства, защищающие ДНК от деградации и слияний конец в конец. Они представлены теломера-capping белком, который распознает теломерную ДНК и регулирует теломеры-формирующий энзим telomerase. Такие capping белки охарактеризованы у ресничатых протозоа. Baumann и Cech описали capping белок, который защищает теломерную ДНК во всем царстве эукариот.
У делящихся дрожжей Schizosaccharomyces pombe авт. идентифицировали открытую рамку считывания с 40% сходства на ее N-конце с теломер-связывающей -субъединицей из hypotrichous ciliate Oxytricha nova. Затем подтвержено, что этот белок, названный Pot1, 'protection of telomeres', связывается специфически с однонитчатой теломерной ДНК. Сродство к ДНК С-терминально укороченного Pot1 было на порядок величин выше, чем сроддство белка полной длины; сходные результаты получены с субъедницей Oxytricha.
Baumann и Cech спорулировали гетерозиготы диплоидной pot1+/pot1- линии. Споры pot1- формировали очень маленькие колонии по сравнению с их pot1+ сестрами, а ДНК окрашивание выявило, что это обусловлено хромосомной нестабильностью - в большинстве pot1- клеток, хромосомы сегрегируют неправильно. Показано, что ДНК из pot1- клеток, которая длиной около 5 kb теряется из теломерной ДНК, подтверждая тем самым, что Pot1 защищает теломерную ДНК.
У человека открыта комплементарная Pot1 ДНК, которая снова обнаруживает высоко законсервированные последовательности вблизи N конца. Клонировали POT1 человека, в результате было подтверждено, что белковый продукт связывается с теломерной ДНК человека. Дрожжевой и человеческий белок наиболее строго связывается со своей собственной теломерной ДНК.
Baumann и Cech определили, что мРНК POT1 присутствует во всех исследованных тканях и показали, что она принадлежит к продуктам гена домашнего хозяйства, "необходима для гаранти интеграции хромосомных концов всех клеток". Установлено также, что Pot1 участвует и в регуляции доступа теломеразы или др. энзимов к концам хромосом.
ORIGINAL RESEARCH PAPERS Pennock, E., Buckley, K. & Lundblad, V. Cell 104, 387-396 (2001) | | |
Chandra, A. et al. Cdc13 both positively and negatively regulates telomere replication. Genes Dev. 15, 404-414 (2001) | | |
Чтобы выживать клетка должна сохранять теломеры. Нужно защищать теломеры от аппарата репарации ДНК - которыый м. помимо прочего рассматривать их как двунитчатые разырвы - т.к. уверен, что они полностью реплицировались во время клеточного деления. Vicki Lundblad и др. показали, что эти две функции обеспечиваются у дрожжей посредством теломер-связывающего белка .
Ранее было показано, что Cdc13 м. позитивно регулировать репликацию теломер, привлекая ответственный энзим - telomerase - к хромосомным концам. Но Cdc13 кроме того негативно модулирует репликацию теломер - эффект, который происходит после рекрутирования теломеразы, и зависимт от белка, известного как .
Объяснением этой негативной регуляции является рекрутирование с помощью Cdc13 активности, защищающей концы. Авт. сливали ДНК-связывающий домен Cdc13 с Stn1. Экспрессия этой конструкции устраняет летальность cdc13 нулевой линии, указывая тем самым на то, что Stn1 является арбитром в защите концов и что он поставляется в теломеры с помощью Cdc13.
Ассоциация Cdc13 как с теломеразой, так и с Stn1 блокируется одной и той же мутацией (cdc13-2), что позволяет предположить, как Cdc13 м. регулировать репликацию теломер. Согласно модели авт., telomerase высвобождается на концах ДНК первой (позитивная ступень). Затем во второй (негативной) ступени Stn1 соединяется с перекрывающим сайтом на Cdc13, это позволяет ему, в свою очередь, быть присоединенным к теломере.
ORIGINAL RESEARCH PAPERS Verdun, R. E. & Karlseder, J. The DNA damage machinery and homologous recombination pathway act consecutively to protect human telomeres. Cell 127, 709–720 (2006)
Геномная стабильность поддерживается благодаря тому, что повреждения ДНК останавливают репликационную вилку и запускают процесс репарации, в то время как теломеры, как полагалось, защищают концы хромосом от запуска подобных реакций. Однако, новое исследование показало, что теломеры действительно нуждаются в damage machinery (кухне повреждений) во время двух самостоятельных моментов клеточного цикла.
Некоторые из сообщений указывают на то, что кухня повреждений ДНК необходима для репликации теломер, так Verdun и Karlseder более детально исследовали время репликации и белки, которые ассоциируют с линиями клеток человека. Они использовали chromatin immunoprecipitation (ChIP) теломерных белков в сочетании с BrdU методом, чтобы показать, что репликация происходит в двух точках клеточного цикла. Дальнейший ChIP анализ показал, что разные сигналы повреждений ДНК запускаются теломерами в каждой из этих точек: в первой остановка репликационной вилки распознается как повреждение ДНК с помощью (ataxia telangiectasia and Rad3 related)-зависимой damage machinery; во второй точке тупые концы реплицирующихся теломер распознаются как разрывы двойной нити и запускается кухня гомологичной рекомбинации с помощью (ataxia telangiectasia mutated)-зависимой передачи сигналов.
Авт. использовали in vitro подход, чтобы показать, что кухня гомологичной репарации необходима для продукции защитных петель, которые характерны для зрелых функциональных теломер. Они поэтому предложили двухступенчатую модель процессинга теломер, согласно которой скорее, чем защита от распознанных повреждений, ДНК damage machinery играет существенную роль.