The multiple nuclear functions of BRCA1: transcription,
ubiquitination and DNA repair
Lea M Starita and Jeffrey D Parvin (jparvin@rics.bwh.harvard.edu) Current Opinion in Cell Biology 2003, 15:345–350
Interest in BRCA1 stems from its role as a tumour suppressor in
breast and ovarian cancer. Intensive research in BRCA1 has
revealed little about its specific role in cancer; rather, this protein
has been implicated in a multitude of important cellular
processes. The diverse biochemical activities of BRCA1
combine to protect the genome from damage. New data reveal
that BRCA1 transcriptionally regulates some DNA-repair genes,
and, in addition, new roles for BRCA1 have been identified in
heterochromatin formation on the X chromosome, doublestrand-
break repair, and ubiquitination. These diverse activities
of BRCA1 may be linked in a single pathway, or BRCA1 might
function in multiple nuclear processes.
Feng, L. et al. MERIT40 facilitates BRCA1 localization and DNA damage repair. Genes Dev. 23, 719–728 (2009)
Wang, B. et al. NBA1, a new player in the Brca1 A complex, is required for DNA damage resistance and checkpoint control. Genes Dev. 23, 729–739 (2009)
Shao, G. et al. MERIT40 controls BRCA1–Rap80 complex integrity and recruitment to DNA double-strand breaks. Genes Dev. 23, 740–754 (2009)
The breast cancer susceptibility gene 1 (BRCA1) формирует комплексы с др. белковыми компонентами, чтобы регулировать клеточную реакцию на повреждения ДНК. Сообщается об идентификации нового компонента BRCA1-содержащего комплекса, который является интегральным для его стабильности и необходим для резистентности к повреждениям ДНК и для G2-M checkpoint контроля.
Используя affinity purification подход, Feng et al. and Shao et al. идентифицировали белок, который они назвали MERIT40 (mediator of RAP80 interactions and targeting 40 kDa), который находится в BRCA1 в комплексе с RAP80 (который содержит ubiquitin-связывающие мотивы), CCDC98 (a coiled-coil домен-содержащий белок, также известный как FAM175A или Abraxas), BRCC36 (deubiquitylating энзим) и BRE (адапторный белок, также известный как BRCC45). Wang et al. также идентифицировали этот белок, который они назвали NBA1 (new component of the BRCA1 A complex), благодаря скринингу на резистентность к ионизирующим облучениям (IR), и показали. что он существует в том же самом BRCA1-содержащем комплексе.
Каждое из трех исследований сообщает конфликтующие данные, согласно которым компонент комплекса BRCA1 белок наз. MERIT40 или NBA1 взаимодействует, общим же мнением является то, что MERIT40/NBA1 существенен для целостности комплекса. Компоненты комплекса, как полагают, независимы и может быть связаны кооперативно с CCDC98, который взаимодействует со всеми членами комплекса. В свою очередь стабильный комплекс необходим для локализации комплекса - и, следовательно, рекрутирование BRCA1 - на места разрывов ДНК. Деплеция любого не-BRCA1 компонента нарушает формирование ДНК повреждениями индуцированных фокусов BRCA1. Напротив, истощение BRCA1 не влияет на локализацию др. компонентов комплекса, это указывает на то, что его функции ниже, чем др. компонентов.
Три группы продемонстрировали, что истощение MERIT40/NBA1 ведет к нарушению G2-M checkpoint контроля и увеличивает чувствительность к IR. BRCA1-содержащие комплексы, как известно, рекрутируются на polyubiquitylated хроматин double-strand breaks (DSBs) вследствие повреждений ДНК. Истощение RAP80 ведет к уменьшению количеств MERIT40/NBA1 на DSBs, это привело Shao et al. к выводу. что рекрутирование BRCA1 комплекса на места повреждений ДНК может обеспечиваться с помощью RAP80. Эти авт. также показали, что deubiquitylating активность BRCC36 существенна для реакций как G2-M checkpoint, так и резистентности к IR.
Биоинформационный анализ, проведенный Wang et al. показал, что BRCA1-содержащий комплекс напоминает комплекс колпачка 26S протеосом. Они также показали, что in vitro несколько др. компонентов BRCA1-содержащих комплексов (помимо RAP80) обладают способностью связывать цепочки polyubiquitin. Хотя функциональное значение этого наблюдения ещё предстоит определить, но полученные данные указывают на то, что MERIT40/NBA1 обеспечивает целостность BRCA1 комплекса, распознавание DSB и активности ubiquitin цепочек в ответ на повреждения ДНК.
Когда опухолевый супрессор BRCA1 мутантен в клетках грудных желез или яичников, то возникают опухоли, тогда как мутации BRCA1 в клетках др. типов летальны.
Неожиданное повсеместное функционирование BRCA1 в ядерных процессах транскрипции, ремоделирования хроматина и молчания и в различных ДНК репарирующих механизмах. Он кроме того обладает активностью ubiquitin ligase. Дальнейшую неразбериху создают ряд белков, которые взаимодействуют с BRCA1. Будут рассмотрены ядерные функции BRCA1, более сложные внеядерные функции BRCA1, включая регуляцию репликации центросом и цитокинез [1-4], обсуждаться не будут в данном обзоре.
BRCA1 and transcription
Концепция BRCA1 как транскрипционного фактора была впервые предположена в подходах по временной трансфекции, показавших, что репортёрный ген активируется с помощью С-конца BRCA1, слитого с GAL4 ДНК-связывающим доменом [5,6]. Но так как такой подход м. давать позитивные результаты и для нетранскрипционных факторов, то функция BRCA1 была подтверждена с помощью ассоциации BRCA1 с РНК полимеразой II в большом комплексе, названном RNA polymerase II holoenzyme (holo-pol) [7]. BRCA1 , как было установлено, регулирует транскрипцию в очищенной системе in vitro [8]. BRCA1 не согласуется с моделью белка, связывающего энхансер, т.к. он не соединяется с ДНК сиквенс-специфически. BRCA1, однако, соединяется с ДНК независимо от последовательностей, но предпочтительно с аномальными структурами [9], это больше всего согласуется с ролью в репарации ДНК скорее. нежели с в транскрипции.
BRCA1 ассоциирует с holo-pol посредством крайних концов:
N-терминальный RING-finger домен обеспечивает первичное связывание с holo-pol, возможно посредством его ассоциации с BARD1 (BRCA1-associated RING domain
protein 1), который также является компонентом holo-pol
[10]. С-конец BRCA1 соединяется также с holo-pol посредством его ассоциации с RNA helicase A [11]. Внутренняя часть BRCA1 соединяется с большим количеством энхансер-связывающих факторов (ряд взаимодействующих транскрипционных факторов показан на Рис. 1), а наиболее преходящие результаты трансфекции согласуются с функцией ко-активатора, в которой BRCA1 перекидывает мостик от специфического энхансер-связывающего фактора к комплексу holo-pol.
Обнаружение генов-мишеней, регулируемых с помощью избыточной экспрессии BRCA1 в культуре ткани м. пролить свет на пути аномальной экспрессии в клетках, которые мутантны по BRCA1. Некоторые эксперименты проведены с использованием технологии микрочипов [12-17]. Чувствительный к p53 ингибитор клеточного цикла и фактор, чувствительный к стрессам, такие как p21 и GADD45, соотв., стимулируются с помощью избыточной экспрессии BRCA1, a BRCA1, как было установлено, является ко-активатором для p53 [12,18-20]. В недавнем исследовании [15] p53 стабилизировался с помощью избыточной экспрессии BRCA1; однако, в отличие от стабилизации p53 с помощью повреждений ДНК BRCA1 стимулировал те p53 пути, которые вели к жизнеспособности и репарации скорее, чем к апоптозу. Напр., BRCA1 синергично стимулировал арест клеточного цикла и
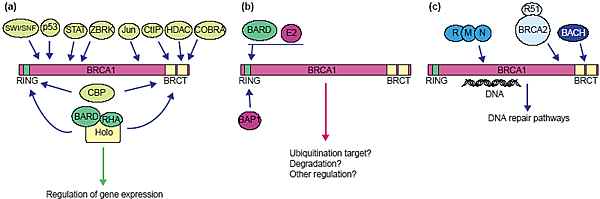
A subset of known nuclear interactions by BRCA1 are shown and grouped according to function: (a) Transcription and chromatin remodeling,
(b) ubiquitination, and (c) DNA repair. The approximate positions of the interaction sites along the BRCA1 polypeptide are indicated by arrows. Some
of the interactions shown are not specifically discussed in this review, but described in other reviews [54]. BARD, BARD1; CBP, CREB-binding protein;
COBRA, cofactor of BRCA1; CtIP, carboxy-terminal binding protein interacting protein; HDAC, histone deacetylase; R51, RAD51; RHA, RNA
helicase A; R-M-N, RAD50-MRE11-NBS1 complex; ZBRK, zinc-finger- and BRCA1-interacting protein with a KRAB domain.
репарацию ДНК повреждений генов, обычно активируемых с помощью p53 (таких как p21 и p53R2). Напротив, PIG3, PERP и Bax гены, участвующие в p53-зависимом апоптозе, подавлялись с помощью избыточной экспрессии BRCA1 [15]. Нокдаун экспрессии BRCA1 с использованием антисмысловой РНК приводит к повышению чувствительности клеток к апоптозу [15].
Интересно, что BRCA1 не всегда защищает клетки от неминуемой гибели - избыточная экспрессия BRCA1 ведет к активации генов, необходимых для апоптоза посредством пути интерферона-γ [16].
BRCA1 and chromatin remodeling
BRCA1 безусловно участвует в транскрипции. Он регулирует многие типы генов, взаимодействуя с репрессорами и активаторами транскрипции и является также частью транскрипционной кухни (machinery). Предполагается, что BRCA1 контролирует структуру хроматина. Как было установлено, он взаимодействует с факторами ремоделирования хроматина SWI/SNF и BRG1 [21,22] и с histone deacetylase [23] (Рис. 1a). Недавно было установлено, что полной длины BRCA1 прикрепляется к гетерохроматину и вызывает частичную декомпрессию ДНК [24]. При таком подходе фрагмент BRCA1, содержащий С-терминальный BRCT (BRCA1 carboxy-terminal repeat) домен значительно ослабляет (reversed) компрессию гетерохроматина. Т.к. удаление домена BRCT даёт гиперактивный белок, то предполагается, что эта функция ремоделирования негативно регулируется с помощью остальной части белка BRCA1 [24]. Хотя эти эксперименты чётко демонстрируют, что BRCA1 играет роль в регуляции поддержания гетерохроматина, природа связывания высоких концентраций белка на amegabase гетерохроматинового домена и декомпрессия с помощью BRCA1 м.б. специфичной для использованной экспериментальной системы.
Побочным продуктом этой находки было наблюдение роли BRCA1 в образовании гетерохроматина. Мутация или потеря функции BRCA1 приводили к изменению фенотипа инактивации Х хромосомы [25], процесса, с помощью которого большой гетерохроматиновый домен образуется по всех Х хромосоме. XIST является молекулой РНК, которая покрывает инактивированную Х хромосому в клетках самок и является центральным для процесса, с помощью которого репрессируется вся хромосома. Если BRCA1 отсутствует в клетке. то XIST РНК не способна локализоваться на Х хромосоме. Присутствие или отсутствие функционального BRCA1 не влияет на уровень транскриптов XIST, а на их локализацию и эффективность в обеспечении молчания. Др. гетерохроматиновые маркёры, такие как макро-гистоны H2A1 и H3, метилированные по Lys9, также не способны локализоваться на неактивной Х хромосоме без функционального BRCA1 [25]. Как же нарушение фенотипа локализации XIST РНК м. способствовать раку груди? Вообще-то де-репрессированная Х хромосома экспрессирует онкогены на более высоком уровне, чем в клетках с одной инактивированной Х хромосомой.
BRCA1 and DNA damage repair
BRCA1 ко-локализуется с macro-H2A1 и H3mK9 (гистоном H3, метилированным по Lys9) на неактивной X; он также ко-локализуется с др. модифицированными гистонами, фосфорилированным H2AX (γ-H2AX), после повреждения ДНК [26]. Нефосфорилированный H2AX разбросам по всему хроматину генома и соответствует повреждениям ДНК, одним из самым ранних событий является фосфорилирование Ser139 в H2AX в больших доменах ДНК, соответствующих миллионам пар оснований [27]. γ-H2AX формирует дискретные фокусы в течение 10 мин. после повреждения ДНК, при этом BRCA1 обнаруживается в этих фокусах. по крайней мере, в течение 30 мин в некоторых линиях клеток и спустя 6 ч в др. После BRCA1 или RAD50 или RAD51, но не оба, также ко-локализуются в фокусах индуцированных повреждений ДНК [26].
Хотя эти фокусы собираются в местах повреждений ДНК замедленность их образования не согласуется со временем основной массы репаративных процессов [28]. Не удивительно, что H2AX-нулевые клетки чрезвычайно чувствительны к ионизирующей радиации и обнаруживают повышенный уровень спонтанных хромосомных аберраций. Формирование BRCA1 и RAD51 индуцированных повреждениями ДНК фокусов
в них уменьшается [29,30], указывая тем самым, что γ-H2AX располагается выше микроскопически обнаруживаемой ассоциации BRCA1 с этими сайтами повреждений. Интересно прикрепление BRCA1 к хромосоме, вызывающее фосфорилирование γ-H2AX, для ко-локализации с BRCA1 без повреждения ДНК. BRCA1 м. , следовательно, рекрутировать киназу, которая фосфорилирует H2AX на ДНК и даёт начало фокусам репарации [24]. Всё это скорее всего указывает на то, что BRCA1 при низких концентрациях рекрутирует H2AX kinase, создавая домены репарации, которые затем собирается в виде фокусов, которые действительно накапливают высокие концентрации BRCA1.
Помимо этих курьёзных фокусов репарации BRCA1 регулирует разнообразные пути репарации повреждений ДНК. BRCA1-дефицитные клетки имеют дефекты по transcription-coupled репарации, гомологичной рекомбинации, nonhomologous end-joining (NHEJ)и microhomology соединению концов [31-36]. In vitro также установлено, что
BRCA1 соединяется с несколькими факторами, участвующими в путях репарации (Рис. 1c). Разнообразие репаративных механизмов указывает на непрямой эффект BRCA1 на эти разнообразные пути. BRCA1 м. траскрипционно активировать гены, кодирующие эффекторы репарации или, напротив, некоторые др. активности, такие как ubiquitination , м. регулировать эффекторные белки. Однако, очевидна биохимическая потребность в нормальной функции BRCA1 для NHEJ в бесклеточных подходах [35], которые скорее всего зависят от ассоциации BRCA1 с комплексом, содержащим RAD50-MRE11-NBS1 сочетание [37,38].
BRCA1 and ubiquitination
Домен RING-finger, такой как тот, что найден в N-конце BRCA1, обычно ассоциирует с активностью ubiquitin ligase. Убиквитин-лигаза полимеризует убиквитин на белке-мишени. Эта убиквитинированная мишень затем деградируется с помощью протеосом. Этот процесс довольно специфичен, т.к. индивидуальные полипептиды в белковом комплексе м.б. убиквитинированы и деградировать, то это м. препятствовать взаимодействию др. интактных субъединиц [39]. BRCA1, вместе с BARD1,
формирует гетеродимер, ассоциирующий посредством RING-finger доменов и соседних спиралей, a BRCA1-BARD1 является активной ubiquitin polymerase (Рис. 1b) [40,41]. Убиквитин полимераза д. синтезировать длинные цепи убиквитина, но ключевым событием д.б. идентификация мишени для действия ubiquitin ligase. Хотя убиквитинирование обычно связано с деградацией белка посредством 26S протеосом,
BRCA1-BARD1 м. также monoubiquitinate белки или полимеризовать убиквитин посредством др. лизиновых связей, в результате такие белки не будут направляться на деградацию, а на некоторые др. цели. BRCA1-BARD1 monoubiquitinates гистоновые мономеры, включая нефосфорилированный H2AX [42,43,44].
Большинство RING-finger белков не специфицируют мишени ubiquitination посредством своих RING-finger доменов, а скорее посредством некоторых др. доменов на том же полипептиде или др. субъединице мультисубъединичной ubiquitin ligases. BRCA1-BARD1 являются крупными белками и поэтому скорее всего, что не тестированные 90% этих белков регулируют ферментативную активность ubiquitin ligase или специфически нацеливают её. Было бы идеальным использовать полные длины BRCA1-BARD1 при специфическом целенаправленном убиквитинировании и идентификации функциональных последствий.
RING пальчики в BRCA1 и BARD1, которые обычно гетеродимеризуются, м. формировать 30 nm кольцеобразные суперструктуры, которые видни в электронный микроскоп. Эти super-RING структуры ферментативно более активны, чем димерные BRCA1-BARD1, и кольцевые структуры функционируют как поддержки (scaffold) для др. белков, чтобы объединить (couple) реакции. E2 ubiquitin conjugating энзим, UbcH5c, формирует кольца, окружающие комплекс BRCA1-BARD1, а цепочки ubiquitin образуют точки на поверхности. surface. Такая организованная поддержка обеспечивает ubiquitination активность BRCA1-
BARD1 на очень высоком уровне [45].
Потенциальная связь этой ubiquitination активности с транскрипционной функцией BRCA1, по-видимому, осуществляется посредством недавно выявленной убиквитинации при активации транскрипции [46]. BRCA1 м. ubiquitinate энхансер-связывающие факторы, ко-активируя их. Или напротив, транскрипционное молчание м. обусловливаться за счёт monoubiquitination гистонов.
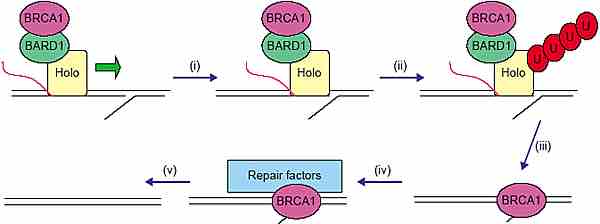
A model is outlined, which links the multiple biochemical activities of BRCA1 into a genome maintenance pathway. BRCA1, along with an elongating
polymerase, would function in the surveillance for DNA damage. Once a damage site is encountered (step 1), BRCA1 would ubiquitinate the
holo-pol, marking it for destruction (step 2), BRCA1 remains bound to the DNA at the lesion (step 3) to recruit the repair factors (step 4), and the lesion is repaired (step 5).
An integrated model for BRCA1 function
in genome stability
BRCA1 участвует в нескольких процессах, но неясно. как это связано с потребностью в BRCA1 во всех типах клеток. Сходным образом с супрессором опухолей p53, BRCA1 активирует гены, кодирующие реакцию репарации ДНК. В отличие от p53, BRCA1 оказывает также непосредственное влияние на процесс репарации. Мы подносили модель, предложенную ранее [47]. Согласно этой модели BRCA1-BARD1 функционирует, надзирая за геномом путём сканирования активных генов в ассоциации с holo-pol, и когда удлиняющих транскрипционный комплекс наталкивается на повреждение ДНК, то BRCA1 инициирует репаративную реакцию (Рис. 2). Интересно отметить, что BRCA1-связывающий кофактор, COBRA1, который регулирует функцию BRCA1 в декомпрессии хроматина [24], как было установлено, необходимая субъединица комплекса, который регулирует элонгацию транскрипции [48]. Когда обнаруживается повреждение на матрице ДНК, то повреждение м.б. скорректировано с помощью transcription-coupled репарации (ступень 1), известная функция BRCA1 [31,32]. Или при некоторых типах повреждений м. потребоваться, чтобы полимераза была удалена для эффективной репарации. Т.к. полимераза, синтезирующая РНК на ДНК-матрице, связана довольно стабильно [49], то мы полагаем, что BRCA1 м. затем ubiquitinate полимеразу, направляя её на деградацию (ступень 2). Хотя имеющихся данных недостаточно для подтверждения участия BRCA1 в этом процессе, но полимераза убиквитинируется и деградирует при повреждениях ДНК [50,51]. Остальная часть BRCA1 комплекса м. оставаться связанной с повреждением ДНК. BRCA1, как установлено. соединяется с ДНК cruciforms и с three-way соединениями, которые м. возникать в местах повреждений [9] (ступень 3). Этот связанный BRCA1 м. затем рекрутировать факторы репарации, такие как RAD50-содержащий комплекс, который м. затем устранять повреждения (ступени 4 и 5).
На это указывает рекрутирование H2AX kinase к сайтам, в которых BRCA1 связан с ДНК [24], и такой надзор за матрицей при транскрипции будет приводить к BRCA1-зависимой деградации транскрипционного аппарата и рекрутированию H2AX kinase, чтобы дать начало сборке репаративных фокусов. Хотя у дрожжей неизвестны гомологи BRCA1, но вообще-то аналогичный путь законсервирован и у них, он обеспечивается с помощью фактора элонгации транскриптов, который генетически связан в этом пути с holo-pol компонентами
[52,53].
Conclusions
Currently, the key cellular functions assigned to BRCA1
are uncomfortably numerous. BRCA1 can interact with
many cellular proteins and pathways, but how these many
interactions address the key questions of required ubiquitous
function and tumour suppressing breast and
ovarian cell function are unclear. We suggest that several
of these pathways can be assimilated into a single pathway,
which controls genomic stability in all cell types.