Пользователи: 
WAVE КОМПЛЕКС
Роль в развитии головного мозга
The WAVE complex in developmental and adulthood brain disorders Kim, HG., Berdasco, C., Nairn, A.C. et al.
 Exp Mol Med (2025). https://doi.org/10.1038/s12276-024-01386-w
|
Actin polymerization and depolymerization are fundamental cellular processes required not only for the embryonic and postnatal development of the brain but also for the maintenance of neuronal plasticity and survival in the adult and aging brain. The orchestrated organization of actin filaments is controlled by various actin regulatory proteins. Wiskott-Aldrich syndrome protein-family verprolin-homologous protein (WAVE) members are key activators of ARP2/3 complex-mediated actin polymerization. WAVE proteins exist as heteropentameric complexes together with regulatory proteins, including CYFIP, NCKAP, ABI and BRK1. The activity of the WAVE complex is tightly regulated by extracellular cues and intracellular signaling to execute its roles in specific intracellular events in brain cells. Notably, dysregulation of the WAVE complex and WAVE complex-mediated cellular processes confers vulnerability to a variety of brain disorders. De novo mutations in WAVE genes and other components of the WAVE complex have been identified in patients with developmental disorders such as intellectual disability, epileptic seizures, schizophrenia, and/or autism spectrum disorder. In addition, alterations in the WAVE complex are implicated in the pathophysiology of Alzheimer’s disease and Parkinson’s disease, as well as in behavioral adaptations to psychostimulants or maladaptive feeding.

|
Полимеризация актина - это АТФ-зависимый спонтанный процесс, который может быть инициирован активацией комплекса 2/3 белков, связанных с актином (ARP)1,2. Лимитирующим шагом в запуске полимеризации актина является образование димеров или тримеров актина. Комплекс ARP2/3 состоит из семи субъединиц, а активированные субъединицы ARP2 и 3 имитируют димер актина, тем самым инициируя полимеризацию актина de novo. Поскольку комплекс ARP2/3 может связываться с уже существующим нитевидным актином (F-актином), он может инициировать образование разветвленной формы F-актина2,3. В мозге разветвленный F-актин играет важную роль в нейронах, включая закрепление синаптических везикул в пресинаптических терминалях, вызывая динамические морфологические изменения в дендритных отростках и связывая постсинаптические белки. Примечательно, что удлинение отростков в олигодендроцитах требует ARP2/3-зависимой полимеризации актина, а разборка актина является необходимым условием для миелинизации аксона4. Динамические морфологические изменения в астроцитах и микроглии зависят от ARP2/3-опосредованной полимеризацией актина5,6.
Комплекс ARP2/3 активируется факторами, способствующими нуклеации 2. Белок синдрома Вискотта-Олдрича (WASP) и верпролин-гомологичный белок семейства WASP (WAVE) относятся к семейству WASP, которые являются наиболее хорошо изученными факторами, способствующими зарождению, и состоят из белков WASP/N-WASP, Scar/WAVE1-3, WASH, JMY, WHAMM и WHIMP 7,8. Три паралога WAVE (WAVE1-3; названия генов: Wasf1, Wasf2 и Wasf3, соответственно) преимущественно, но не исключительно экспрессируются в различных типах клеток мозга (рис. 1 и табл. 1). WAVE1 - это регулятор, обогащающий мозг, который особенно часто встречается в нейронах 9-12, клетках-предшественниках олигодендроцитов и зрелых олигодендроцитах 13. WAVE2 экспрессируется на высоком уровне в микроглии и эндотелиальных клетках сосудов 14,15. WAVE3 также экспрессируется в основном в мозге 16, где его уровень в интернейронах и астроцитах относительно высок 17,18. Белки WAVE существуют в виде гетеропентамерных белковых комплексов, называемых WAVE-комплексом 19,20 или WAVE-регуляторным комплексом 21, вместе с NCK-ассоциированным белком 1 (NCKAP1) или NCK-ассоциированным белком 1-like (NCKAP1L), цитоплазматическим FMR1 взаимодействующим белком (CYFIP) 1 или 2, ABL-интерактором (ABI) 1, 2 или 3, и BRK1 (также называемым HSPC300) 11,19,22,23. По сравнению со своими паралогами, WAVE1, CYFIP2, NCKAP1 и ABI2 относительно многочисленны в головном мозге 19,20,22 (рис. 1). WASP-гомологичный 2 (WH2)-центрально-кислотный (WCA) домен WAVE связывается с комплексом ARP2/3 и активирует его 24,25, в то время как CYFIP, NCKAP, ABI и BRK1 участвуют в регуляции формирования комплекса WAVE, его стабильности, субклеточной локализации или связывания с лигандами или активаторами выше по течению 21,26,27,28. Как и белки WAVE, паралогичные члены каждой субъединицы экспрессируются преимущественно, но не исключительно в определенных типах клеток мозга (рис. 1 и табл. 1). Вероятно, существуют promiscuous (разносторонние) взаимодействия между различными паралогичными субъединицами пентамерного комплекса, что увеличивает регуляторное и функциональное разнообразие, хотя конкретного правила для взаимодействия между паралогичными субъединицами не установлено.
 Fig. 1: Expression pattern of subunit paralogs of the WAVE complex. a Scatter plot of single-cell RNA-seq data for single cells isolated from the whole hippocampus and cortex (left panel). The glutamatergic, GABAergic, and non-neuronal cell types corresponding to the segregated cells are indicated (right panel). Publicly available single-cell RNA-seq data for WAVE genes: Wasf1, 2 and 3 (b), Abi1, 2 and 3 (c), Cyfip1 and 2 (d), Nckap1 and 1l (e) and Brk1 (f). Allen Institute for Brain Science (https://portal.brain-map.org/atlases-and-data/rnaseq).
Table 1 Relative expression of the subunit paralogs of the WAVE complex in brain cell types. The WAVE complex is a cellular activity-dependent actin regulator
Множество сигнальных путей сходятся на комплексе WAVE, что приводит к динамической регуляции полимеризации актина (рис. 2). Первоначально WAVE был идентифицирован как нисходящая мишень Rac, ГТФазы семейства Rho, и как медиатор Rac-индуцированной реорганизации актинового цитоскелета 25,29. Rac1 связывается с двумя сайтами на CYFIP в комплексе WAVE, что приводит к активации WAVE 28,30-32. Протеомный анализ показал, что WRP, Rac-селективный GTPase-активирующий белок, связывается с WAVE1 через свой SH3-домен (Src homology domain 3) и подавляет способность Rac регулировать WAVE1-опосредованную полимеризацию актина 33. Кроме того, ARF1 напрямую связывается с отдельным сайтом на CYFIP1, который значительно усиливается при связывании RAC1, и таким образом активирует комплекс WAVE 34,35. Кислые фосфолипиды, такие как фосфатидилинозитол (3,4,5)-трифосфат (PIP 3) или PI(4,5)P 2, связываются с основной поверхностью комплексов WAVE2, усиливая активацию комплекса WAVE2 через пренилированный Rac1 20 (рис. 2). Однако на активацию нативного комплекса WAVE1, выделенного из мозга крупного рогатого скота, не влияли пренилированные Rac1 или PIP 2, что указывает на потенциальные различия в регуляции комплексов WAVE1 и WAVE2 20.
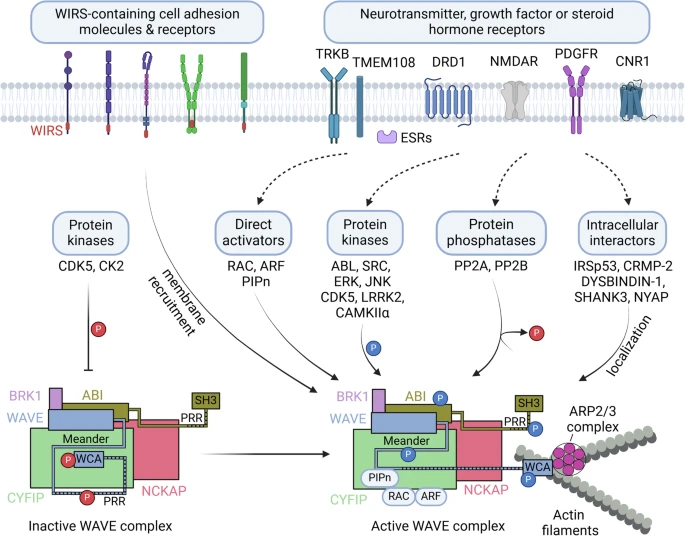 Fig. 2: A proposed model for various input signals and regulators converging on the WAVE complex, leading to ARP2/3 complex-mediated actin polymerization. Constitutively active kinases (CDK5 and CK2) basally phosphorylate WAVE, and the intramolecular interaction of the WCA domain with the meander region in WAVE inhibits actin polymerization. In contrast, activated neurotransmitter, growth factor or steroid hormone receptors and their signaling pathways may activate various regulators of the WAVE complex, including direct activators (such as RAC, ARF and PIPn), protein kinases or phosphatases that modulate phosphorylation levels in the WAVE complex, and protein interactors that localize the WAVE complex at locally active sites. WIRS-containing cell adhesion molecules or receptors recruit the WAVE complex to cell membranes. The integration of such input signals results in spatially and temporally orchestrated ARP2/3-mediated actin polymerization in a variety of cellular events. This figure was created with BioRender.com.
Фосфорилирование комплекса WAVE - еще один важный регуляторный механизм. Фосфорилирование WAVE или ABI опосредует активацию или ингибирование комплекса WAVE, в зависимости от конкретных сайтов фосфорилирования 22,36 (рис. 2 и 3). Доступность WCA-области WAVE для активации ARP2/3 затруднена из-за многочисленных взаимодействий как с CYFIP, так и с “meander” областью WAVE 21 (рис. 2). Тирозинкиназа ABL фосфорилирует Tyr151 из WAVE1, Tyr150 из WAVE2 или Tyr151 из WAVE3 37,38, и это фосфорилирование, как предполагается, препятствует взаимодействию между областью “meander” WAVE и CYFIP, освобождая область WCA для активации с помощью ARP2/321 (рис. 2 и 3a-c). Не-рецепторная тирозинкиназа SRC также фосфорилирует WAVE1 по Tyr125, что, как предполагается, дестабилизирует “meander” область WAVE и высвобождает область WCA 21,39. Фосфорилирование WAVE2 по Ser137 циклин-зависимой киназе 5 (CDK5) в области “meander” необходимо для PDGF-индуцированной миграции клеток-предшественников олигодендроцитов in vitro40. ABL также фосфорилирует дополнительные Tyr-сайты, включая остатки 248, 337 и 486 WAVE3 37,38. WAVE2 фосфорилируется киназой, регулируемой внеклеточным сигналом (ERK), или Jun N-terminal kinase (JNK) на Ser308, Ser343, Thr346 и Ser351, и это фосфорилирование необходимо для полимеризации актина и миграции культивируемых клеточных линий 41-43. Однако эти сайты фосфорилирования ERK или JNK отсутствуют в WAVE1 и WAVE3 36. Фосфорилирование ABL, SRC и CDK5 в области “meander” и фосфорилирование ERK в богатой пролином области опосредуют активацию WAVE, необходимую для формирования ламеллиподий и миграции клеток 36, однако роль фосфорилирования в этих участках в функционировании комплекса WAVE в мозге еще предстоит выяснить.
 Fig. 3: Regulation of the WAVE complex by phosphorylation. Phosphorylation at multiple sites on WAVE (a–c) and ABI1 (d) is regulated by protein kinases or protein phosphatases. The proposed roles of phosphorylation (stimulatory as blue, inhibitory as red, and uncharacterized as black) are indicated. The phosphorylation sites are illustrated for human sequences. WHD WAVE homology domain, B basic domain, PRR proline-rich region, WCA WASP homology 2 (WH2), central, acidic domain, WAB WAVE-binding domain, SNARE soluble N-ethylmaleimide-sensitive factor-activating protein receptor domain, HHR Hox homology region, SR serine/threonine-rich region, PP polyproline region, SH3 Src homology 3 domain.
Комплекс WAVE1 был идентифицирован из лизатов головного мозга как интерактор CDK5 в комплексе с p35, активируемый субъединицей CDK522. CDK5 фосфорилирует Ser310, Ser397 и Ser441 в пролин-богатой области WAVE1, но эти три сайта не фосфорилируются ERK. Фосфорилирование CDK5 нативного комплекса WAVE1, очищенного из мозга крысы, или рекомбинантного WAVE1 ингибирует ARP2/3-опосредованную полимеризацию актина in vitro22. Ser310 является основным сайтом фосфорилирования, а мутация Ser310Ala блокирует ингибирующий эффект фосфорилирования CDK5 на WAVE1-опосредованную полимеризацию актина in vitro22. По сравнению с белком WT экспрессия мутанта Ser310Ala в нейронах с нокдауном WAVE1 оказывала большее влияние на восстановление морфологии дендритных отростков, тогда как экспрессия фосфомиметического мутанта Ser310Asp практически не влияла на фенотип, вызванный нокдауном WAVE122. Примечательно, что CDK5 фосфорилирует WAVE1 на Ser310, Ser397 и Ser441 с высокой базальной стехиометрией в интактном мозге, срезах мозга или в культивируемых нейронах22. При стимуляции рецепторов дофамина D1 (DRD1) или глутаматных рецепторов NMDA в срезах мозга сигналы цАМФ и кальция и нижележащие протеинфосфатазы-2A (PP2A) и PP2B дефосфорилируют WAVE1, что указывает на критическое участие протеинфосфатаз в активации WAVE1, вызванной активностью нейронов44 (рис. 2 и 3а). В первичных культивируемых кортикальных нейронах повторная деполяризация KCl вызывает дефосфорилирование WAVE1 в зависимости от NMDA-рецепторов45. В культивируемых нейронах коры головного мозга крысы фосфорилирование WAVE1 по Ser310, Ser397 и Ser441 резко увеличивалось под действием эстрогенов46. Повышенное фосфорилирование и привлечение CDK5 и WAVE1 к клеточной мембране ассоциировалось с увеличением толщины актина и мембранных выпячиваний. Интересно, что изменения в фосфорилировании WAVE1, опосредованном CDK5, были предложены в качестве последующего события RAC и G-белковой сигнализации46. Более того, изменения фосфорилирования WAVE1 под действием CDK5 наблюдались в мозге мышиной модели болезни Альцгеймера (AD)47, а также в мозге мышей, которым вводили кокаин11.
Казеиновая киназа 2 (CK2) фосфорилирует Ser482, 484, 488, 489 и 497 в кислой области WCA-домена WAVE248. Первые три сериновых участка консервативны для всех WAVE: Ser489 характерен для WAVE2, а Ser497 в WAVE2 сохраняется и в WAVE3 (Ser501)36 (рис. 3a-c). CK2 - конститутивно активная киназа, которая фосфорилирует сайты в кислых областях. На основании исследования, проведенного в Dictyostelium, предполагается, что фосфорилирование первых двух сайтов CK2 (WAVE1-Ser544 и 546, WAVE2-Ser482 и 484 и WAVE3-Ser487 и 489) ингибирует функцию WCA через облегчение внутримолекулярных взаимодействий с основной областью в WAVE49. Таким образом, фосфорилирование CK2 домена WCA WAVEs и/или фосфорилирование CDK5 пролин-богатой области WAVE1 может приводить к ингибиторному состоянию, в то время как дефосфорилирование этих участков, опосредованное протеинфосфатазами, может активировать комплекс WAVE22,43,44,48,49.
ABI обогащен в дендритных отростках и локализуется в постсинаптической плотности (PSD)50. Ca2+/кальмодулин-зависимая киназа IIα (CAMKIIα) взаимодействует с ABI1 и фосфорилирует Ser88, что может быть необходимо для ABI-опосредованного созревания дендритных отростков51 (рис. 3д). ERK также фосфорилирует множество сайтов в ABI1, включая Ser183, Ser216, Ser225, Thr265, Ser267, Ser392, Thr394 и Ser41041 (рис. 3д). Эксперименты с белками, содержащими мутации этих сайтов, указывают на роль фосфорилирования ABI1 в высвобождении домена WCA и последующей активации комплекса WAVE36,41. Примечательно, что сайты фосфорилирования, киназы и функциональные последствия могут варьировать в зависимости от типа клеток и клеточных условий20,41. Более того, значимость CK2- или ERK-опосредованных механизмов фосфорилирования в мозге или клетках мозга еще предстоит выяснить. Другие паралоги субъединиц, включая ABI220, ABI352,53 и CYFIP1/254, также являются фосфопротеинами, но киназы для конкретных сайтов фосфорилирования и роль их фосфорилирования в активности комплекса WAVE или полимеризации актина еще предстоит выяснить.
Транслокация и мембранный рекрутинг комплекса WAVE критически важны для вовлечения WAVE-опосредованной полимеризации актина в специфических функциональных локусах. Помимо RAC (или ARF) и кислых фосфолипидов (PIP2 или PIP3), многие трансмембранные молекулы клеточной адгезии и рецепторы, включая протокадхерины, ROBO, нетриновые рецепторы, нейролигины, GPCRs и ионные каналы, могут рекрутировать комплекс WAVE на мембраны через взаимодействие CYFIP и ABI с консервативным мотивом WIRS (WAVE complex interacting receptor sequence)23,28 (рис. 2). Например, у C. elegans SYG-1, молекула адгезии, взаимодействует с комплексом WAVE через мотив WIRS, и это взаимодействие необходимо для динамики синаптического F-актина, формирования синапса и ветвления аксона55. Некоторые другие партнеры по связыванию WAVE1, такие как collapsin response mediator protein 2 (CRMP или DPYSL2)56 или каннабиноидный CB1-рецептор (CNR1)57, WAVE2 взаимодействует с субстратом инсулинового рецептора 53, IRSp53 (BAIAP2) или дистробревин-связывающим белком-1 (DYSBINDIN-1 или DTNBP1), Взаимодействие CYFIP с retrolinkin (TMEM108)58 и связывание ABI1 с PSD-белком SHANK350 также способствуют оркестровке активности комплекса WAVE локально во время аксонального и дендритного роста или синаптического ремоделирования (рис. 2).
The WAVE complex mediates various cellular events in the brain
Synapse formation and plasticity
F-актин высоко обогащен в дендритных отростках, постсинаптических частях возбуждающих синапсов 59 и пресинаптических терминалях 60, и роль комплекса WAVE в этих синаптических компартментах была подробно рассмотрена 23 (рис. 4а). Комплекс WAVE1 локализован в дендритных отростках и играет важную роль в регуляции морфологии дендритных отростков. Делеция WAVE1 уменьшает плотность и изменяет морфологию дендритных отростков 22,61. Экспериментальное снижение экспрессии WAVE1 приводит к уменьшению плотности зрелых дендритных отростков с более крупной головкой и короткой шейкой, но увеличивает плотность филоподиальных дендритных выпячиваний 22. WAVE1, эффектор нисходящего потока сигнализации NMDA-рецепторов 44, является важнейшим медиатором когнитивных функций. У мышей с нокдауном GluN1 (GluN1KD) снижен уровень NMDA-рецепторов, наблюдается дефицит плотности стриатумных dendritic spines и когнитивные нарушения. У мышей с нокаутом GluN1KD снижены уровни компонентов пути RAC1 в стриатуме, причем дефицит RAC1 и WAVE1 наблюдается в возрасте 6 и 12 недель. Одновременно с этим у мышей этих возрастов наблюдается дефицит плотности dendritic spines (отростков) у средних колючих нейронов (MSN). Скрещивание мышей GluN1KD с мышами, экспрессирующими WAVE1, позволило восстановить уровень белка WAVE1 в стриатуме и MSN spine density, а также поведение в тестах Y-maze и 8-плечевом радиальном лабиринте 62. WAVE2 также регулирует плотность и морфологию MSN spine вместе с IRSp5363 или DYSBINDIN-164, а ABI1 взаимодействует с богатой пролином областью SHANK3 через свой SH3-домен и регулирует формирование синапсов 50. Поскольку генетические варианты DYSBINDIN-1 связаны с шизофренией, взаимодействие DYSBINDIN-1 с WAVE2 и ABI1 и их участие в регуляции дендритных отростков может иметь отношение к патофизиологии шизофрении 50,64.
 Fig. 4: The WAVE complex mediates various cellular events, and alterations in the WAVE complex are associated with brain disorders. a Subunit proteins in the WAVE complex are known to be involved in specific cellular events. b Alterations (blue box) in specific subunit proteins in the WAVE complex are associated with the pathophysiology (yellow box) of NDDs, AD, PD, cocaine reward and binge eating behavior. This figure was created with BioRender.com.
CYFIP1 и 2 обогащены в дендритных отростках возбуждающих синапсов и участвуют в поддержании сложности дендритных структур и дендритных отростков65,66. Увеличение числа копий CYFIP1 связано с изменениями в нейральном развитии. Избыточно экспрессирующие CYFIP1 нейроны и трансгенные мыши, сверхэкспрессирующие CYFIP1, демонстрируют незрелые дендритные отростки67,68. CYFIP1 и 2 также экспрессируются в тормозных синапсах67. Избыточная экспрессия CYFIP1 в культивируемых нейронах гиппокампа увеличивает частоту mEPSC и уменьшает амплитуду mIPSC. Напротив, делеция CYFIP1 в возбуждающих нейронах у мышей повышает уровень нейролигина 3 и субъединиц GABAA-рецепторов β2/3 и увеличивает тормозную постсинаптическую кластеризацию и mIPSC67. Дисбаланс возбуждающей и тормозной синаптической функции, вероятно, способствует патофизиологии мутаций CYFIP или вариаций числа копий, связанных с аутизмом и SCZ.
Локальная трансляция мРНК в дендритах нейронов важна для синаптической пластичности и развития мозга. CYFIP1 напрямую связывается с рибонуклеопротеином 1 (FMR1 или FMRP) и опосредует репрессивную активность FMRP в трансляции дендритных белков. CYFIP1/Sra1 напрямую связывается с eIF4E (эукариотическим фактором инициации трансляции 4E) и тем самым ингибирует инициацию трансляции69. Важно отметить, что BDNF или DHPG, агонист метаботропных глутаматных рецепторов группы I, вызывают диссоциацию CYFIP1 и eIF4E, что указывает на ключевую роль комплекса FMRP-CYFIP1 в индуцированном активностью нейронов локальном синтезе белка69. Таким образом, CYFIP1 участвует в различных функциях FMRP-CYFIP1-eIF4E и WAVE-комплекса и тем самым координирует локальную трансляцию белка и полимеризацию актина в дендритных отростках66,69,70. Стимуляция BDNF высвобождает CYFIP1 из eIF4E через активацию RAC1, но стабилизирует и активирует CYFIP1 в комплексе WAVE66. Таким образом, путь BDNF-RAC1-CYFIP1 может увеличивать как локальную трансляцию белков, так и полимеризацию актина, необходимую для синаптической пластичности66,69.
Neurite and axonal outgrowth
WAVE1 также регулирует динамику конуса роста и рост нейритов в культивируемых нейронах61 (рис. 4а). Было установлено, что WAVE1 вместе с RAC GEF Dock3 являются эффекторами нижнего течения сигнального каскада BDNF-TRKB во время аксонального роста71. Retrolinkin является регулятором эндоцитарной транспортировки комплекса BDNF-TRKB. Retrolinkin взаимодействует с CYFIP2 в комплексе WAVE1 и способствует BDNF-индуцированному эндоцитозу TRKB и аксональному росту58. Комплекс WAVE1 взаимодействует с белками семейства NYAP (Neuronal tYrosine-phosphorylated Adaptor for the PI 3-kinase) и участвует в PI3K-опосредованном росте нейритов и морфогенезе нейронов72. CRMP-2 играет важную роль в росте аксонов и спецификации аксон-дендритов. CRMP-2 взаимодействует как с комплексом WAVE1, так и с кинезином-1. CRMP-2 и кинезин-1 необходимы для локализации WAVE1 в конусах роста, а WAVE1 и CYFIP1 - для CRMP-2-опосредованного роста аксона и спецификации аксон-дендритов56. CYFIP1 играет роль в определении размера пресинаптических окончаний и вероятности высвобождения везикул во время развития73. CYFIP2 регулирует пресинаптические функции, предположительно, через регуляцию количества пресинаптических митохондрий во взрослом мозге74.
Neurogenesis and neuronal migration
CYFIP1 необходим для активности нейральных стволовых клеток в субвентрикулярной зоне, для поддержания нейрогенной ниши боковых желудочков и, таким образом, нейрогенеза коры головного мозга у взрослых мышей75 (рис. 4а). CYFIP1 также регулирует нейрогенез гиппокампа у взрослых через клеточно-автономные и не клеточно-автономные механизмы76. Гаплонедостаточность мышиного Cyfip1 приводит к увеличению числа нейронов гиппокампа взрослых за счет снижения апоптоза без изменения пролиферации вследствие неспособности микроглии индуцировать апоптоз путем секреции соответствующих факторов76. Кроме того, гаплонедостаточность Cyfip1 вызывает аномальную миграцию взрослых нейронов из-за изменения ARP2/3-опосредованной динамики актина76. Комплекс WAVE2-ABI2 участвует в радиальной миграции нейронов, направляемой глиями77. Комплекс WAVE2-ABI2, контролирующий активность конусов роста, имеет решающее значение для завершения перехода от мультиполярной к биполярной форме до начала направляемой глией миграции. Активность конуса роста регулируется ABL и CDK5 путем фосфорилирования Tyr150 и Ser137 WAVE2. Таким образом, комплекс WAVE2-ABI2 важен для определения конечного местоположения мигрирующих нейронов в неокортексе.
Oligodendrocyte morphogenesis and axonal myelination
Опосредованная комплексом ARP2/3 полимеризация актина важна для начальной фазы разрастания олигодендроцитов и закрепощения аксонов, но быстрая обмотка миелина инициируется разборкой F-актина4,78. WAVE1 высоко экспрессируется в олигодендроцитах13. По сравнению с олигодендроцитами WT, первичные культивированные олигодендроциты, выделенные из мышей с нокаутом WAVE1 (KO), демонстрируют меньшее количество отростков. Гипомиелинизация в мозолистом теле и дефект образования миелина в зрительных нервах in vivo также наблюдались у нокаутных мышей WAVE1, что ясно указывает на роль WAVE1 в развитии олигодендроцитов и миелинизации13 (рис. 4а). Ген CYFIP1 является кандидатом в факторы риска психопатологии, связанной с вариантом числа копий (CNV) 15q11.2 BP1-BP2. У крыс и мышей с гаплонедостаточностью Cyfip1 наблюдались обширные изменения белого вещества, включая истончение миелиновой оболочки в мозолистом теле, что позволяет предположить потенциальный вклад дефектов миелинизации в повышенный риск психических расстройств, наблюдаемый у людей с делецией числа копий 15q11.279,80.
Mitochondrial function and trafficking
WAVE1 был идентифицирован как компонент комплекса, связанного с митохондриями и содержащего PKA, протеинфосфатазу-1, проапоптотический белок BAD и глюкокиназу в печени81. Иммуноэлектронная микроскопия выявила локализацию WAVE1 на внешней мембране митохондрий81. Кроме того, WAVE1 был идентифицирован в ходе анализа белков, связывающих киназы (AKAPs), и охарактеризован как мультикиназный комплекс формирования каркаса, включающий PKA или ABL82. В головном мозге фокальный ишемический инсульт вызывает образование белкового комплекса, состоящего из WAVE1, pancortin-2 и антиапоптотического белка Bcl-xL. Этот белковый комплекс связан с митохондриями и рекрутирует Bax, который участвует в апоптозе, вызванном ишемическим инсультом83. В первичных культивируемых нейронах WAVE1-опосредованная полимеризация актина участвует в индуцированном нейронной активностью движении и делении митохондрий45 (рис. 4а). Поскольку функция митохондрий критически важна для развития нейронов, формирования и пластичности синапсов, WAVE1-опосредованные изменения митохондрий могут способствовать WAVE1-зависимым синаптическим изменениям.
Nuclear function
Обработка клеток leptomycin B (ингибитором ядерного экспортера CRM-1) вызывает ядерное накопление ABI184 и CYFIP285. При применении NMDA ABI1 транслоцируется из PSD в ядра первичных культивируемых нейронов гиппокампа и пирамидных нейронов на свежих срезах мозга крысы50. В ядре ABI1 взаимодействует с комплексом транскрипционных факторов белков Myc/Max и усиливает транскрипцию генов, регулируемых E-box50. Исследования Xenopus и Drosophila также дают представление о ядерной функции комплекса WAVE. Ядерный WAVE1 был идентифицирован как центральный регулятор транскрипционного перепрограммирования, необходимого для развития ооцита Xenopus86. WAVE1 присутствует в ядре ооцита. Нокдаун WAVE1 в эмбрионах приводит к дефектной активации генов Hox и аномальному развитию. Роль ядерного WAVE1 в активации генов во время развития, вероятно, опосредована доменом гомологии WAVE (WHD), что указывает на потенциальное участие других субъединиц комплекса WAVE в ядерном программировании86. WAVE, Scar и ARP2/3-зависимая полимеризация ядерного актина и миозина у дрозофилы также критична для восстановления и стабильности гетерохроматина87. Примечательно, что специфические ядерные события и транскрипция генов регулируются наличием мономерного актина, поскольку различные ядерные белки, такие как комплексы ремоделирования хроматина88, модификаторы гистонов89 и РНК-полимеразы90,91, требуют мономерного актина для своей активности. Напротив, при разрыве двойных нитей ДНК ядерный F-актин необходим для работы путей репарации с не-гомологичным концевым соединением и гомологичной рекомбинацией92. Таким образом, опосредованная комплексом WAVE полимеризация актина может контролировать доступность глобулярного актина (G-актина) или F-актина, которые необходимы для ядерного программирования или репарации ДНК. Роль ядерных путей, опосредованных комплексом WAVE, в развитии, функционировании и нарушениях мозга еще предстоит изучить (рис. 4а).
The WAVE complex in neurodevelopmental disorders (NDDs)
WASF1 (WAVE1 gene) variants
Дисфункция WAVE1 ассоциируется с различными NDDs, представляя собой широкий фенотипический спектр от легкой до тяжелой умственной отсталости (ID), расстройств аутистического спектра (ASD), эпилепсии и задержки развития ( DD). Потеря WAVE1 приводит к сенсомоторной задержке и снижению способностей к обучению и памяти у мышей 10. Секвенирование нового поколения выявило десять различных вариантов последовательности в WASF1. Ito и др. впервые идентифицировали три de novo гетерозиготных усеченных варианта, сгруппированных вокруг С-концевого домена WH2, у пяти не-родственных индивидуумов с перекрывающимися фенотипами нейроразвития, включая тяжелое нарушение интеллекта с аутистическими чертами и судорогами 93 (табл. 2а). Среди двух нонсенс-вариантов и одного варианта со сдвигом рамки считывания рекуррентный вариант p.Arg506Ter был идентифицирован у трех человек 94-97. Другой нонсенс-вариант, p.Gln520Ter, также был зарегистрирован у одного человека 98,99. Для вариантов, усекающих белок WAVE1, авторы предполагают усиление функции или доминантно-отрицательную функцию из-за присутствия усеченного белка в клетках пораженных индивидуумов 93. Примечательно, что эти три усеченных варианта ассоциируются с фенотипом, сходным с тем, который наблюдается у мышей с нокаутом Wasf110. В связи с тем, что в базе данных Genome Aggregation Database (gnomAD) от здоровых людей 100 зарегистрировано мало вариантов с потерей функции (LoF) - всего 17 случаев (частота аллелей: ~7E-7), ген WASF1 демонстрирует исключительно высокую непереносимость таких мутаций. Кроме того, оценка вероятности непереносимости LoF (pLI) 0,91 для WASF1 93 говорит о том, что LoF, скорее всего, лежит в основе механизма этого варианта. Сообщалось еще о двух предполагаемых вариантах LoF. Гетерозиготная делеция, обусловленная CNV, охватывающей экзоны 8-10 WASF1 и включающей весь ген GPR6, была выявлена у монозиготных близнецов с ID, аутизмом и эпилепсией 95 (табл. 2b). Внутригенная гетерозиготная делеция, охватывающая экзоны 4-11 гена WASF1, была обнаружена у человека с психоневрологическим расстройством 101 (табл. 2c).
Table 2 Alterations of the WAVE complex in brain disorders.
Остальные пять гетерозиготных миссенс-вариантов были зарегистрированы у пациентов с NDDs (табл. 2d). Фенотипы двух независимых пациентов, каждый из которых несет вариант p.Trp161Arg в WASF1, возникший в результате различных геномных изменений - либо c.481T > A95, либо c.481T > C102, - схожи и проявляются в виде ID, DD, эпилепсии и микроцефалии. Рекуррентные варианты p.Leu455Pro и p.Leu535Pro были обнаружены у пациента с аутизмом и у другого пациента с DD, соответственно103,104. Кроме того, вариант p.Trp161Cys был обнаружен у пациента с ID аутизмом, гипотонией и эпилепсией, а p.Lys172Glu - у пациента с DD, гипотонией и аутизмом95. Помимо выраженной непереносимости усеченных вариантов, WASF1 с оценкой миссенса Z score 2,99 также предсказывается как непереносимый к миссенс-вариантам. Никаких заметных фенотипических различий между LoF-вариантами и миссенс-вариантами не наблюдалось, это указывает на то, что мутировавшие остатки расположены в функционально важных доменах. Trp161 и Lys172 находятся в WHD (также называемом WH1, WASP homology 1, доменом) и базовой области, соответственно95, в то время как Leu535 расположен в домене WH2. В базе данных ClinVar были указаны еще четыре нонсенс-варианта, ассоциированных с НDD: p.Lys46Ter, p.Glu88Ter, p.Met165AsnfsTer8 и p.Ser489Ter (табл. 2e). Кроме того, включен один миссенс-вариант, p.Ser440Leu, для которого нет фенотипической информации (табл. 2f). Интересно, что микроцефалия, которая не наблюдается у нокаутных мышей, проявилась у двух монозиготных близнецов с делецией, охватывающей весь ген GPR6 и экзоны 8-10 WASF195, а также у двух независимых пациентов, несущих идентичный миссенс-вариант p.Trp161Arg95,102. Однако этот фенотип не наблюдался ни у одного другого пациента с мутацией WASF1.
WASF2 variants
WAVE2 экспрессируется на относительно высоком уровне в микроглии, периваскулярных макрофагах и сосудистых эндотелиальных клетках мозга (рис. 1б и табл. 1). Два миссенс-варианта WASF2, c.113G > A (p.Arg38Gln) и c.494T > C (p.Leu165Pro), были выявлены у пациентов с аутизмом и NDDs103-105, однако функциональные изменения, связанные с нарушениями мозга, остаются неизученными (табл. 2g).
WASF3 variants
Хотя WAVE3 экспрессируется в нейронных клетках, в частности в GABAергических интернейронах (рис. 1b и табл. 1), его функция менее хорошо изучена, чем функция WAVE1. Нонсенс-вариант WASF3, p.Glu88Ter, был идентифицирован у пациента с НDD103,104,106 (табл. 2h).
CYFIP1 variants
CYFIP1 был связан с аутизмом107,108. CNVs в области 15q11.2 встречаются у пациентов с NDDs включая ASD, SCZ, ID и эпилепсию (табл. 2i). Самый маленький регион, связанный с этим состоянием, находится между точками разрыва BP1 и BP2 на хромосоме 15q11.2. Среди четырех генов (TUBGCP5, CYFIP1, NIPA2 и NIPA1), включенных в этот узкий регион, все больше доказательств указывают на значимость CYFIP1109.
Вслед за ассоциацией двух общих SNP в CYFIP1 у пациентов с SCZ110 (табл. 2j), с помощью секвенирования экзома были выявлены сложные гетерозиготные варианты p.Ile476Val и p.Pro742Leu у двух больных мужчин из одной семьи, у которых наблюдались DD, ID, ASD, эпилепсия и отсутствие речи (табл. 2k). Эти мутации происходят в белковых доменах, отвечающих за поддержание взаимодействий внутри комплекса WAVE. Трансгенные модели мух с этими мутациями демонстрируют аномальную морфологию нейронов и потерю F-актина, что отражает основные поведенческие симптомы, такие как дефицит социального взаимодействия и двигательной координации. Молекулярный и клеточный анализ фибробластов кожи одного из пробандов выявил дефицит полимеризации актина, что, возможно, связано с нарушением функциональности комплекса WAVE и снижением стабильности CYFIP1 в составе комплекса111.
Дисрегуляция синтеза белка в нейронах была вовлечена в патогенез различных НDD112. В подгруппе пациентов с синдромом хрупкой Х (FXS), у которых наблюдаются тяжелые формы аутизма и обсессивно-компульсивного поведения, мРНК CYFIP1 снижена, особенно у пациентов с фенотипом Прадера-Вилли108. Снижение регуляции CYFIP1 или генетические изменения в CYFIP1 могут изменять FMRP-CYFIP1-eIF4E-опосредованное подавление локального синтеза белка в нейронах69. У пациентов с ASD были выявлены четыре гетерозиготных миссенс-варианта CYFIP1, включая распространенный вариант, p.Gly820Ser113 (Таблица 2l), и три редких варианта, p.Asp544Asn, p.Met956Arg и p.Ala1025Thr114 (Таблица 2m). Кроме того, у пациента с диагнозом SCZ115 был выявлен полигенный паттерн, включающий редкий гетерозиготный деструктивный нонсенс-вариант, p.Arg624Ter, в CYFIP1 (табл. 2n). Кроме того, у пациента с синдромом West, который характеризуется инфантильной ID эпилепсией и ASD116, была выявлена внутри-кадровая делеция аминокислоты, p.Val305del (табл. 2o).
CYFIP2 (PIR121) variants
Как и CYFIP1, CYFIP2 присутствует как в возбуждающих, так и в тормозных синапсах, где взаимодействует как с комплексом WAVE, так и с FMRP117. Три de novo гетерозиготных миссенс-варианта в CYFIP2 были выявлены у четырех не-родственных людей с диагнозом ранней эпилептической энцефалопатии с помощью трио полноэкзомного секвенирования (WES), что позволяет предположить ассоциацию этого гена с НDD. Все три варианта затрагивают Arg87 - p.Arg87Cys, p.Arg87Leu и p.Arg87Pro - что позволяет предположить, что этот ген представляет собой «горячую точку» для этих мутаций (табл. 2p). Сопутствующие фенотипы включают тяжелую психомоторную задержку, ID, черепно-лицевые аномалии и гипотонию118. Интересно, что синдром West был диагностирован у двух человек118, как и у пациента с вариантом CYFIP1116. Основываясь на оценке непереносимости LoF (pLI = 1) для CYFIP2, авторы первоначально предположили, что варианты в Arg87 могут приводить к нарушению WAVE-опосредованной динамики актина через механизм LoF, но последующее белковое моделирование и функциональный анализ предполагают модель устойчивой активации комплекса WAVE вариантами Arg87118. Однако механизм LoF CYFIP2 был дополнительно подтвержден обнаружением нестабильности белка вариантов CYFIP2 в клетках119 и в мозге120. Мышиная модель knock-in варианта p.Arg87Cys рекапитулировала многие неврологические и нейроповеденческие фенотипы, встречающиеся у пациентов с этим вариантом120. Кроме того, у мышей knock-in наблюдалась дезорганизация слоев нейронов и глиоз в мозге. Вариант Arg87Cys усиливает убиквитинирование и протеасомную деградацию CYFIP2 в культивируемых клетках и мозге мышей, устанавливая механизм LoF с участием вариантов Arg87120. С помощью трио секвенирования нового поколения было выявлено восемь различных вариантов de novo в CYFIP2 у 12 независимых пациентов, которые имели фенотип, характеризующийся ID, судорогами и мышечной гипотонией. Один из них является сплайс-вариантом, а остальные семь - миссенс-вариантами (табл. 2q). Эти варианты пространственно группируются в третичной структуре и, по прогнозам, ослабляют взаимодействие с WAVE1 или NCKAP1 в комплексе WAVE121.
Гомозиготная делеция Cyfip2 приводит к летальному исходу у мышей, что указывает на жизненно важную роль CYFIP2 в раннем развитии, в то время как гетерозиготные нокаутные мыши демонстрируют аномальное поведение и кортикальные дендритные шипы, аналогичные тем, что наблюдаются у Fmr1-нулевых мышей122. Поэтому, наряду с оценкой LoF непереносимости CYFIP2 (pLI = 1), авторы первоначально предположили, что гаплонедостаточность CYFIP2 вызывает фенотип нарушения нейрального развития у человека. Однако фенотип в гетерозиготной мышиной модели Cyfip2 KO был более мягким, чем тяжелый фенотип, наблюдаемый у людей, гетерозиготных по варианту121. В генах, связанных с аутосомно-доминантными заболеваниями, гетерозиготных мутаций достаточно для проявления фенотипа у человека. Для сравнения, гетерозиготные нокаутные мыши не проявляют явного фенотипа, в то время как гомозиготные нокаутные мыши проявляют фенотип, сходный с человеческим123. Далее Zweier et al. предположили, что обнаруженные варианты способствуют частичному LoF CYFIP2, особенно пять миссенс-вариантов (p.Arg87Cys, p.Ile664Met, p.Glu665Lys, p.Asp724His и p. Gln725Arg), которые расположены вблизи интерфейса CYFIP2-WAVE1 и, как предполагается, нарушают связывание WCA-области WAVE и высвобождение WCA для ARP2/3-опосредованной полимеризации актина, что предполагает сценарий усиления функции121.
Begemannи др. сообщили еще о 19 людях, имеющих варианты CYFIP2, благодаря секвенированию экзома, в том числе о 14 новых вариантах (табл. 2r). У пяти человек было зарегистрировано два варианта, p.Arg87Cys и p.Asp724His, причем p.Asp724 был вторым мутационным очагом. Среди них три человека, имеющие варианты LoF - p.Lys501Ter, p.Ser258Glnfs2 и c.2058-1G > C - проявляли такие клинические признаки, как DD, ID синдром дефицита внимания и гиперактивности (ADHD), поведенческие проблемы, судороги, аномалии черепно-лицевой области и цифровые аномалии. Фенотипы людей с миссенс-вариантами включали DD, ID, эпилепсию, гипсаритмию, микроцефалию, лицевой дисморфизм, дисфагию, проблемы со зрением и нарушения сна. Авторы утверждали, что у пациентов с вариантами LoF фенотипы не столь тяжелы, как у людей с миссенс-вариантами124. Однако CYFIP2 нетерпим как к миссенс-вариантам (z = 6,01), так и к LoF-вариантам (pLI = 1). Кроме того, патогенные варианты в одном и том же гене нейроразвития могут приводить к различным клиническим проявлениям, включая различия в степени тяжести125. Следовательно, эти мутации, вероятно, приводят к LoF CYFIP2.
NCKAP1 (HEM2) variants
Нонсенс-вариант (p.Glu1100Ter) NCKAP1 был идентифицирован как сегрегационный вариант во многих поколенbz[ семьи, состоящей из 13 членов, страдающих легкой формой ID ADHD, макроцефалией и высоким ростом (табл. 2s). Этот вариант экспрессируется в клетках различных областей мозга, включая клетки Пуркинье, зубчатое ядро мозжечка, область CA4 и зубчатую извилину гиппокампа, а также лобные области серого и белого вещества126.
В коллекции Simons Simplex Collection (SSC) две de novo усекающие мутации в NCKAP1 - p.Gly175TrpfsTer14 и p.Glu1088Ter - были выявлены у двух пробандов с ASD из двух не-родственных семей с симплексным quad127. Анализ данных секвенирования генома 2 308 человек из 493 мультиплексных семей с аутизмом из Autism Genetic Resource Exchange (AGRE), проведенный с помощью мега-анализа TADA (Transmitted And De novo Association), выявил нарушения NCKAP1 среди 69 генов, ассоциированных с риском развития аутизма128.
В 20 не-родственных семьях было зарегистрировано в общей сложности 19 предполагаемых пагубных вариантов в NCKAP1. Эти варианты включают четыре варианта со сдвигом рамки, пять нонсенс-вариантов, пять сплайс-вариантов и три миссенс-варианта (табл. 2t). Кроме того, были выявлены микроделеция de novo размером 240 кб, охватывающая три гена, включая NCKAP1 в 2q32.1, и одна сбалансированная хромосомная инверсия de novo [46,XX,inv(2)(p23.1q32.1)dn], усекающая NCKAP1 в точке разрыва геномной инверсии 2q32.1. Среди них 13 были обнаружены de novo, четыре передавались по наследству, а три имели неизвестный характер наследования129.
NCKAP1L (HEM1) variants
NCKAP1L специфичен для гемопоэтических клеток и экспрессируется избирательно в микроглии в мозге (рис. 1е и табл. 1). Мутации NCKAP1L у людей и делеция HEM1 у мышей вызывают широкую иммунную дисрегуляцию130-132. Секвенирование следующего поколения выявило один интронный114, два миссенс-133,134 и два нонсенс-варианта у больных аутизмом104,114, а функциональные последствия этих вариантов еще предстоит установить (табл. 2u).
ABI1 variants
Нонсенс-вариант, c.367C > T, p.Arg213Ter, в ABI1 был идентифицирован у пациента с сердечно-сосудистой дисфункцией и НDD135,136,137 (табл. 2v). Кроме того, два миссенс-варианта, c.584C > T, p.Thr195Ile, и c.1238A > C, p.Glu413Ala, в ABI1 были обнаружены у пациентов с миеломенингоцеле (spina bifida)138 и ASD104,114, соответственно. Эмбрионы Abi1-нулевых мышей меньше, с некоторыми не-перевернутыми эмбрионами и волнистостью нервной трубки уже в возрасте E8.5139. У некоторых эмбрионов нервная трубка была открыта. Abi1-нулевые мышиные эмбрионы демонстрировали летальность между E10.5 и E11.5139. В клетках Abi1-KO наблюдался дефект регуляции актинового цитоскелета, который объяснялся изменением активности комплексов WAVE1 и 2. По сравнению с контрольными клетками уровни компонентов WAVE-комплекса WAVE1/2, CYFIP1/2 и NCKAP1 были значительно ниже, а уровни белка ABI2 в клетках Abi1-KO были в два раза выше139.
ABI2 variants
Две гетерозиготные миссенс-мутации de novo, ABI2-p.Arg373Cys и p.Glu202Lys, были идентифицированы у двух пробандов ASD из двух независимых семей с симплексным квадрантом на основании данных экзомного секвенирования, полученных из коллекции Simons Simplex Collection (SSC)127 (табл. 2w). Кроме того, гомозиготная мутация LoF, p.Arg132Ter, в ABI2 была выявлена с помощью комбинации генотипирования с помощью микрочипов, картирования гомозиготности по происхождению (HBD), анализа CNV и секвенирования экзома. Это открытие было сделано в когорте из 192 мультиплексных пакистанских и иранских кровосмесительных семей. ABI2 - один из 26 вновь выявленных генов-кандидатов, ассоциированных с аутосомно-рецессивным несиндромальным ID140 (табл. 2).
Гомозиготная делеция мышиного Abi2 приводила к аномальным фенотипам глаз и мозга, которые являются тканями с наиболее высокой экспрессией ABI2. Кроме того, потеря ABI2 привела к дефектам миграции клеток в неокортексе и гиппокампе, а также к нарушениям морфологии и плотности дендритных шипов. Эти нарушения сопровождаются серьезным дефицитом как кратковременной, так и долговременной памяти141.
ABI3 variants
NESH (NEw molecule containing SH3) был первоначально выделен из библиотеки кДНК плаценты человека как новый ген, содержащий SH3-домен, а также богатые пролином и серином области. Из-за сходства с ABI1 и ABI2 он был впоследствии включен в семейство ABI и обозначен как ABI3142. Ниже рассматривается мутация ABI3, связанная с AD (табл. 2y).
BRK1 (HSPC300) variants
Вариант инициирующего кодона c.2T > A, p.Met1? и вариант регуляторной последовательности c. *135T > C в BRK1 были выявлены у двух пациентов с расстройством аутистического спектра114, однако функциональные последствия этих вариантов еще предстоит изучить (табл. 2z).
The role of the WAVE complex in neurodegenerative diseases
AD
Изменения в комплексе WAVE были вовлечены в патофизиологию AD. Снижение экспрессии WASF1 (ген WAVE1 человека) наблюдалось в мозге человека при AD по сравнению с мозгом здорового человека и в мышиных моделях AD по сравнению с контрольными143. Снижение экспрессии WASF1 в мозге при AD также наблюдалось в общедоступных наборах данных144,145. Вероятный механизм, подтвержденный экспериментальными данными, заключается в том, что AICD (внутриклеточный домен APP), образующийся в результате амилоидогенного пути, связывается с промотором WAVE1 и негативно регулирует его транскрипцию и экспрессию белка143. С другой стороны, WAVE1 взаимодействует с APP и колокализуется с ним в аппарате Гольджи. Экспериментальное снижение экспрессии WAVE1 уменьшает отпочкование APP-содержащих везикул из аппарата Гольджи и снижает трафикинг APP к клеточной поверхности, тем самым подавляя продукцию Aβ143. Нокаут WAVE1 резко снижает уровень Аβ и восстанавливает дефицит памяти у трансгенных мышей APP/PS1, экспрессирующих семейные мутации в APP (APPswe) и presenilin1 (PSEN1 ΔE9)146, что свидетельствует о защитной роли сниженной экспрессии WAVE1 при AD143. Ко-локализация белка WAVE1 или фосфорилированного WAVE1 с агрегированным гиперфосфорилированным tau наблюдалась в мозге тройных трансгенных мышей (APPswe/PS1M146V/tauP301L)47 и пациентов с AD147. Ген WAVE1 также был предложен в качестве узлового гена, связанного с AD, на основании исследований мышиных моделей АД148 или мозга человека с AD149.
Уровень RAC1 в лобной коре пациентов с AD снижен по сравнению с контрольной группой без деменции, но он преходяще снижен в мозге животной модели AD (3xTg, 7 месяцев)150. Активация RAC1 в клеточной линии или первичных культивируемых кортикальных нейронах увеличивала уровень Аβ и гиперфосфорилирование tau. Ингибирование RAC1 ослабляло γ-секретазную активность в отношении APP151. Также было обнаружено, что экспрессия NCKAP1 заметно снижена в мозге больных AD по сравнению с нормальными людьми152, однако функциональная роль снижения экспрессии NCKAP1 при AD еще не установлена.
Снижение экспрессии CYFIP2 наблюдалось в посмертном мозге больных AD и в мышиной модели AD153. Уровень CYFIP1 также снижен у людей с поздней стадией AD, но уровни белка как CYFIP1, так и CYFIP2 повышены в посмертных тканях гиппокампа пациентов с фронтотемпоральной лобарной дегенерацией (FTLD) с патологией TDP43154. Несколько сходных с AD патологий, включая накопление Аβ, гиперфосфорилирование tau, глиоз и потерю дендритных спинов в гиппокампе, наблюдались у гетерозиготных мышей с нокаутом Cyfip2153,155, но не у мышей с нейрон-специфическим условным нокаутом Cyfip2156. Исследователи не смогли точно определить, почему нокаут CYFIP2 влияет на AD-подобные патологии, хотя нокаут WAVE1 на амилоидную патологию является защитным143. Эти противоположные результаты могут быть связаны с ингибирующим действием CYFIP2 на WAVE1-опосредованную полимеризацию актина118,157 или с участием CYFIP2 в трансляции мРНК155.
Генетические исследования AD выявили вариант кодирования ABI3, p.Ser209Phe (rs616338), ассоциированный с поздним развитием AD158-161 (табл. 2y). Наличие Phe209 нарушает общее фосфорилирование ABI3, делая белок ABI3 функционально неактивным. Это изменение может быть истолковано как гипоморфная функция или частичная LoF53. ABI3 высоко экспрессируется в микроглии162,163 (рис. 1c и табл. 1), и экспрессия ABI3 повышена в посмертном мозге при AD162,164. Делеция Abi3 у мышей ускоряет накопление Aβ, уменьшает миграцию микроглии и кластеризацию вокруг бляшек, а также снижает LTP из-за накопления Aβ в мышиной модели AD (5XFAD)164,165. Кроме того, нокаут Abi3 также увеличивает массу тела и жира и снижает энергозатраты166. Благодаря генетическому совпадению Abi3 и Gngt2 на хромосоме 11q у мышей (отсутствует у людей на 17q21.32 с направлением обратной транскрипции), нокаут Abi3 также направлен на Gngt253. Двойной нокаут Abi3-Gngt2 предрасполагает мышей к иммунному фенотипу, характеризующемуся реактивным глиозом и экспрессией генов, напоминающих те, которые ассоциируются с AD. Примечательно, что гены Trem2, Plcg2, Tyrobp и Csf1r, ассоциированные с AD, повышаются у этих мышей дозозависимым образом даже в отсутствие накопления Aβ У трансгенных по APP мышей TgCRND8 потеря Abi3-Gngt2 приводит к дозозависимому от гена и возраста снижению отложения Aβ, а у мышей Abi3-Gngt2-/- опосредованная AAV экспрессия способствующей агрегации формы человеческого tau усугубляет тауопатию и астроцитоз53.
Parkinson’s disease (PD)
Киназа 2 с богатыми лейциновыми повторами (LRRK2) является наиболее распространенным геном, связанным с PD. Мутации LRRK2 ассоциируются с семейной и спорадической PD а LRRK2 является перспективной терапевтической мишенью для PD67. На модели дегенерации глазау дрозофилы, вызванной hLRRK2, был проведен скрининг генетических модификаторов LRRK2, и было установлено, что Scar/WAVE является генетическим интерактором LRRK2168. В микроглии BV2 нокдаун LRRK2 снижает уровень WAVE2, но не снижает уровни WAVE1 и WAVE3. Уровень WAVE2 также снижен в первичной микроглии и макрофагах, полученных из костного мозга мышей с нокаутом LRRK2. Однако в микроглии и макрофагах, полученных от нокаутных мышей LRRK2-G2019S, наблюдается увеличение экспрессии WAVE2169. LRRK2 связывает и фосфорилирует мышиный WAVE2 на Thr470 (Thr471 в человеческом WAVE2, рис. 3б), стабилизируя и предотвращая протеасомную деградацию WAVE2, тогда как LRRK2 не взаимодействует с WAVE1169. Макрофаги и микроглия, выделенные от пациентов с PD или мышей с LRRK2-G2019S, демонстрируют повышенное фосфорилирование Thr470 на WAVE2 и усиленный фагоцитарный ответ, а делеция LRRK2 вызывает противоположные эффекты. Кроме того, WAVE2 и его фосфорилирование участвуют в LRRK2-G2019S-индуцированной гибели дофаминергических нейронов в ко-культурах макрофагов и среднего мозга. Кроме того, нокдаун WAVE2 у мышей и нокдаун Scar у дрозофилы ослабляют LRRK2-G2019S-индуцированную гибель дофаминергических нейронов169. Ингибитор киназы LRRK2 или ингибитор ARP2/3 предотвращает активацию микроглии и нейротоксичность in vitro169,170, что позволяет предположить участие пути LRRK2-WAVE2 в микроглии в патогенезе PD
The role of the WAVE complex in other brain disorders
Поведенческие реакции, вызванные психостимуляторами, требуют опосредованной комплексом WAVE1 синаптической модуляции. Субстраты мышей C57BL/6 отличаются по локомоторной активности и сенсибилизированной реакции на кокаин и метамфетамин. Генетический скрининг мышей C57BL/6N, у которых острая и сенсибилизированная реакция на психостимуляторы ниже, чем у мышей C57BL/6J, выявил мутацию Ser968Phe в Cyfip2 в качестве причинного варианта171. Мутация Ser968Phe дестабилизирует CYFIP2, а делеция аллеля у мышей C57BL/6N приводит к фенотипам острого и сенсибилизированного ответа на кокаин, что позволяет предположить, что CYFIP2 является критическим регулятором ответа на кокаин171. В качестве альтернативы, крио-электронная микроскопия комплекса WAVE с RAC1 показала, что мутация Ser968Phe в CYFIP1 (соответствующая Ser968Phe в CYFIP2) расположена непосредственно под сайтом связывания RAC1. Дальнейшая характеристика показала, что Ser968Phe в CYFIP1 способствует связыванию RAC1 и активации WAVE-комплекса172. У мышей с нокаутом CYFIP2 Ser968Phe ослаблена вызванная кокаином поведенческая сенсибилизация и условное предпочтение места173. Вызванное кокаином увеличение экспрессии c-Fos и GluA1, а также увеличение плотности дендритных спинов в nucleus accumbens, ключевой области мозга для вознаграждения кокаином, наблюдалось у мышей дикого типа, но не у мышей с нокаутом CYFIP2 Ser968Phe, это указывает на значимость этого варианта в реакции на кокаин173. В другом исследовании, проведенном на штаммах мышей BXD, было высказано предположение, что Cyfip2 является геном-кандидатом, связанным с различными реакциями на потребление в парадигме самопоглощения кокаина174.
WAVE1 также необходим для поведенческой и синаптической пластичности, вызванной кокаином11. Кокаин блокирует пресинаптическую обратную отдачу дофамина175 и вызывает высвобождение дофамина176, тем самым повышая уровень синаптического дофамина. Делеция WAVE1 в нейронах, экспрессирующих рецептор дофамина D1, но не рецептор дофамина D2, значительно снижает вознаграждение от кокаина, измеряемое по кокаин-ассоциированному предпочтению места и кокаин-индуцированному сенсибилизированному локомоторному поведению11. Введение кокаина также снижает ингибирующее фосфорилирование Ser310, 397 и 441 на WAVE1 в зависимости от рецептора NMDA или рецептора дофамина D1. WAVE1 необходим в нейронах, экспрессирующих рецептор дофамина D1, для вызванной кокаином глутаматергической синаптической пластичности11. В совокупности эти результаты свидетельствуют о критической роли WAVE1 и CYFIP2 в синаптической пластичности в наградных областях мозга, опосредующих краткосрочные и долгосрочные поведенческие реакции на психостимулирующие препараты.
CYFIP2 был также идентифицирован как значимый генетический фактор, лежащий в основе переедания, при анализе близкородственных субстратов мышей C57BL/6177. Мыши C57BL/6N, но не C57BL/6J, демонстрировали быструю и устойчивую эскалацию потребления вкусной пищи. У гетерозиготных нокаутных мышей Cyfip2 на фоне C57BL/6N наблюдалось снижение переедания, сравнимое с таковым у мышей дикого типа C57BL/6J. Поскольку CYFIP1 удален или импринтирован у пациентов с синдромом Прадера-Вилли I типа - НDD, характеризующимся крайней гиперфагией и гиперреактивностью системы вознаграждения на сигналы, связанные с пищей, общие биологические функции CYFIP1 и 2 могут быть связаны с дезадаптивным питанием у людей177,178.
Current limitations and remaining questions
Несмотря на целый ряд убедительных результатов, наше понимание механизмов регуляции комплекса WAVE в конкретных типах клеток мозга остается неполным. Предыдущие исследования установили взаимодействие между кислыми фосфолипидами, белковыми активаторами, мембран-ассоциированными или цитоплазматическими партнерами по связыванию и протеинкиназными или фосфатазными механизмами полной активации комплекса WAVE с помощью анализов восстановления in vitro и подходов структурной биологии (рис. 2). В структурных исследованиях комплекса WAVE in vitro использовался рекомбинантный комплекс WAVE, содержащий mini-WAVE21,28,30,172, в котором отсутствует богатая пролином область из-за неупорядоченности этой области и трудности кристаллизации21. Хотя WAVE1 демонстрирует высокий уровень базального фосфорилирования в нейронах или мозге22,49, это базовое фосфорилирование не присутствует в очищенном рекомбинантном комплексе WAVE. Более того, в нескольких исследованиях использовались интактные комплексы WAVE, выделенные из мозга или клеток19,20,22,179. Нативный полноразмерный комплекс WAVE2 успешно активировался совместным действием пренилированного RAC-GTP, кислых фосфолипидов и фосфорилирования20. Однако интерпретация полимеризации актина in vitro, особенно с использованием нативного полноразмерного комплекса WAVE1, иногда ограничена, а механизм его регуляции все еще не до конца понятен из-за возможных артефактов, наблюдаемых в условиях in vitro, или неспособности воспроизвести активацию нативного комплекса WAVE1 с помощью RAC1 и PIP319,20. Для оценки изменений в комплексе WAVE в мозге или нейронах использовались антитела, специфичные для сайтов фосфорилирования, или измерения уровней экспрессии субъединиц комплекса WAVE. Однако технически невозможно оценить активность комплекса WAVE in vivo, на которую влияют многочисленные регуляторные факторы, включая белковые активаторы, фосфолипиды, фосфорилирование или дефосфорилирование, а также мембран-ассоциированные интеракторы (рис. 2).
Изменения в регуляторных субъединицах комплекса WAVE, вероятно, влияют на WAVE-опосредованную полимеризацию актина. Однако, поскольку ABI может регулировать N-WASP180, CYFIP может связываться с FMRP и eIF4E69, а также с несколькими РНК-связывающими белками181, а NCKAP1L может также взаимодействовать с комплексом mTOR 2 через рапамицин-нечувствительного компаньона mTOR (RICTOR)130, поэтому исследователи не определили окончательно, зависит ли влияние ассоциированных с заболеванием вариантов CYFIP, NCKAP или ABI от WAVE-опосредованной полимеризации актина или не зависит от нее. Таким образом, важно полностью понять, вносит ли повышенная или пониженная полимеризация актина, опосредованная комплексом WAVE, вклад в прогрессирование заболевания.
Доказательств ключевого вклада изменений в комплексе WAVE в развитие НDDs и нейродегенеративных заболеваний становится все больше (рис. 4b). Изменения в WAVE1, ABI1/2 и CYFIP1/2 в нейронах или ABI3 и WAVE2 в микроглии были вовлечены в патофизиологию NDDs AD или PD Потенциальный вклад дисфункции комплекса WAVE в других типах клеток, таких как олигодендроциты и астроциты, в развитие заболевания еще предстоит изучить. Недавние исследования показали, что инфильтрация лейкоцитами и аберрантное воспаление оказывают защитное или пагубное влияние на развитие нейродегенеративных заболеваний182,183. Вопрос о том, вовлечены ли WAVE2, NCKAP1L и CYFIP2 в функции Т-клеток130,184,185; WAVE1 и WAVE2 в функции макрофагов186,187; или WAVE-комплекс-опосредованная полимеризация актина в периферических и центральных иммунных клетках в патогенные или защитные механизмы при заболеваниях мозга, еще предстоит изучить. Интригующий вопрос заключается в том, каким образом усиление или ослабление функции комплекса WAVE сегрегации в качестве фактора риска для, по-видимому, различных заболеваний развития по сравнению со взрослыми или нейродегенеративными заболеваниями (табл. 2 и рис. 4b). Создание in vivo моделей вариантов комплекса WAVE120,173 для характеристики роли функциональных изменений в комплексе WAVE в прогрессировании заболеваний станет важнейшей задачей на ближайшие годы.
|