Ползователи: 
РЕТРОТРАНСПОЗОНЫ LINE-1
Структура, функции и адаптпции
Structures, functions and adaptations of the human LINE-1 ORF2 protein Eric T. Baldwin, Trevor van Eeuwen, David Hoyos, et al.
 Nature volume 626, pages194–206 (2024)
|
The LINE-1 (L1) retrotransposon is an ancient genetic parasite that has written around one-third of the human genome through a ‘copy and paste’ mechanism catalysed by its multifunctional enzyme, open reading frame 2 protein (ORF2p)1. ORF2p reverse transcriptase (RT) and endonuclease activities have been implicated in the pathophysiology of cancer2,3, autoimmunity4,5 and ageing6,7, making ORF2p a potential therapeutic target. However, a lack of structural and mechanistic knowledge has hampered efforts to rationally exploit it. We report structures of the human ORF2p ‘core’ (residues 238–1061, including the RT domain) by X-ray crystallography and cryo-electron microscopy in several conformational states. Our analyses identified two previously undescribed folded domains, extensive contacts to RNA templates and associated adaptations that contribute to unique aspects of the L1 replication cycle. Computed integrative structural models of full-length ORF2p show a dynamic closed-ring conformation that appears to open during retrotransposition. We characterize ORF2p RT inhibition and reveal its underlying structural basis. Imaging and biochemistry show that non-canonical cytosolic ORF2p RT activity can produce RNA:DNA hybrids, activating innate immune signalling through cGAS/STING and resulting in interferon production6,7,8. In contrast to retroviral RTs, L1 RT is efficiently primed by short RNAs and hairpins, which probably explains cytosolic priming. Other biochemical activities including processivity, DNA-directed polymerization, non-templated base addition and template switching together allow us to propose a revised L1 insertion model. Finally, our evolutionary analysis demonstrates structural conservation between ORF2p and other RNA- and DNA-dependent polymerases. We therefore provide key mechanistic insights into L1 polymerization and insertion, shed light on the evolutionary history of L1 and enable rational drug development targeting L1.

|
В последнее время в эволюции транспозонов приматов преобладают РНК-ретротранспозоны, которые вставляют РНК-посредники в геном с помощью активности обратной транскриптазы (RT) 9. Эти ретротранспозоны делятся на два класса: (1) эндогенные ретровирусы (ERVs), фланкированные длинными терминальными повторами (LTRs), и (2) не-LTR ретротранспозоны long interspersed element-1 (LINE-1, L1) 1. Считается, что ERV больше не активны у человека 1. Напротив, каждый человек наследует около 100 полиморфных и фиксированных потенциально активных L1, что составляет небольшое подмножество из примерно полумиллиона неактивных копий и фрагментов L1 11. LINEs ко-эволюционируют со своими хозяевами на протяжении 1-2 миллиардов лет, с момента появления эукариот. Человеческий L1 кодирует два белка, ORF1p 10 и ORF2p, последний из которых обладает эндонуклеазной (EN) и RT активностями 11-13, а также тремя другими доменами с неизвестными функциями (рис. 1a,b). ORF2p котрансляционно связывается с кодирующей его L1 РНК, это называется "цис-преференцией "14-17, образуя рибонуклеопротеиновый (РНП) комплекс с множеством копий ORF1 и белков хозяина 10,15,17-19 (рис. 1б). Новые инсерции начинаются с механизма priming (затравки) с target primed reverse transcription (TPRT): EN-узелок на "нижней" нити ДНК высвобождает 3'-OH ДНК, используемый для priming RT и образования промежуточного гибрида РНК:ДНК 20-23. Детали TPRT в L1, синтеза второй нити и того, как разрешаются образующиеся промежуточные продукты, остаются неясными, хотя известно, что последующий ступенчатый разрыв второй "верхней" нити ДНК 24 приводит к характерной дупликации целевого сайта, обычно не превышающей 20 пар нуклеотидов (п.н.), фланкирующей L1-опосредованные инсерции 24,25. Несмотря на то, что ORF2p предпочитает цис, он также связывает и вставляет другие РНК, включая последовательности мРНК и РНК с короткими промежуточными элементами, такими как Alu.
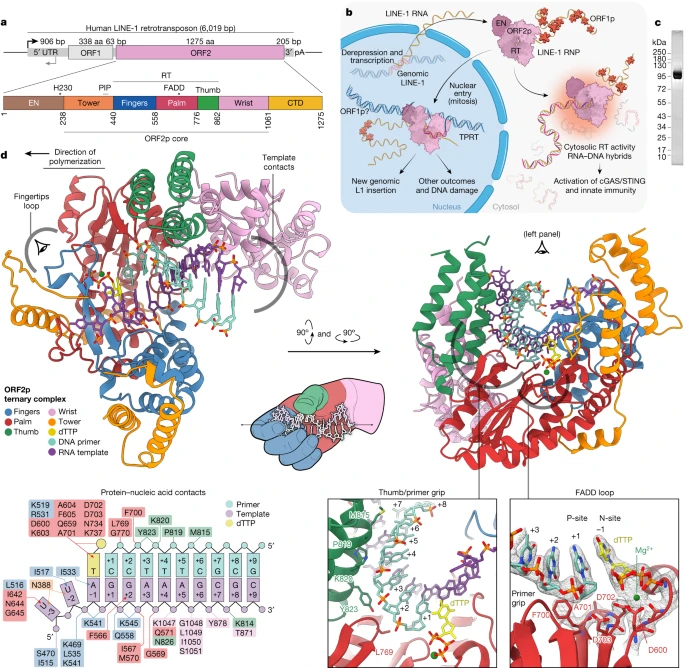 Fig. 1: Pathogenic replication cycle of L1 and the 2.1 A resolution crystal structure of human ORF2p core in a ternary complex.
a, The 6 kb human L1 element contains an internal 5' untranslated region (UTR) promoter, two proteins ORF1p and ORF2p in a bicistronic arrangement separated by 63 nt and a short 3' UTR. b, Replication cycle of L1, a streamlined self-copying DNA parasite. Derepression of genomic L1s results in Pol II transcription and export of the L1 RNA, which is translated to form an RNP complex containing one copy ORF2p, a multifunctional enzyme, and many copies of ORF1p, a homotrimeric chaperone involved in nuclear entry that can form phase-separated granules. Canonically, in the nucleus, ORF2p integrates a new copy of the L1 RNA into the genome in a mechanism termed TPRT, in which cleavage by the L1 EN liberates a genomic DNA (gDNA) 3'-OH used to prime reverse transcription of the L1 RNA, followed by insertion by poorly understood mechanisms (‘Discussion’, Fig. 6). Non-canonical outcomes contribute to pathology: failed insertions and aberrant EN activity result in DNA damage and translocations, and aberrant cytosolic RT activity generates inflammatory RNA:DNA hybrids. Host proteins (not shown) are associated at every step and may repress L1 or function as essential cofactors. c, Sodium dodecyl sulfate polyacrylamide gel electrophoresis analysis of pure, monodisperse 97 kDa ORF2p core after size exclusion chromatography. d, Two new domains (tower and wrist) and three canonical RT subdomains (fingers, palm, thumb) coordinate with a hybrid duplex RNA template (purple) and DNA primer (cyan) and incoming dTTP nucleotide (yellow) for ORF2p core RT activity in the 2.1 A resolution crystal structure in a ‘right-hand’ RT fold that is uniquely adapted. All five ORF2p core domains contact the template or primer, and numerous residues contact the incoming base; protein contacts are summarized in the inset schematic.
Депрессированные элементы L1 могут способствовать развитию патологии рака, старения, нейродегенерации и воспаления (механизмы представлены на рис. 1б). В соответствии с этим ингибиторы RT показали многообещающие результаты в модельных системах6-8,26,27 и в клинических исследованиях колоректального рака28 и синдрома Aicardi–Goutieres, редкой менделевской интерферонопатии, характеризующейся накоплением промежуточных элементов L14,27,29. Однако наши знания о механических деталях как вставки L1, так и того, как L1 вносит вклад в патофизиологию, ограничены. Наиболее хорошо охарактеризованными родственниками L1 являются элементы R2 LINE насекомых21 и бактериальные мобильные интроны группы II30,31, которые не имеют N-концевой апуриновой/апиримидиновой EN (APE)-подобной EN из ORF2p12,13 и дивергировали из человеческой линии около 700 миллионов и 4 миллиардов лет назад, соответственно. Оба распознают и мобилизуют уникальные последовательности ДНК и РНК, что ограничивает возможность сравнения с L1.
Чтобы устранить пробелы в знаниях о биологии L1 и облегчить поиск лекарств, мы создали системы для очистки полноразмерного ORF2p и минимального "ядра", охарактеризовали RT-активность ORF2p и определили его структуру с помощью различных методов. Наши исследования показали: (1) эффективный прайминг RT короткими РНК и шпильками; (2) прямой цитозольный синтез РНК: ДНК-гибридов, которые активируют cGAS-STING, что приводит к выработке интерферона; (3) ряд конформационных адаптаций в "правосторонние" пальцевые, ладонные и большой пальце упаковки RT, которые, вероятно, модулируют биохимические активности, необходимые для цикла репликации L1; (4) наличие двух ранее не описанных доменов в ядре RT, которые мы назвали "башня" и "запястье" (‘tower’ and ‘wrist’); и (5) согласованную динамику N-концевого EN и карбокси-концевого домена (CTD). Опираясь на эту структуру, мы прояснили эволюционные связи между консервативными структурными особенностями ORF2p. Наши результаты проливают свет на ранее загадочные этапы цикла репликации L1, его роль в патофизиологии и потенциальные пути создания терапевтических средств.
Purification of highly active ORF2p RT
Предыдущие попытки измерить ферментативную активность ORF2p были ограничены невозможностью очистить более чем следовые количества ORF2p RT, при этом ограниченные характеристики нечистого фермента указывали на то, что ORF2p может осуществлять синтез ДНК с использованием РНК или ДНК-шаблонов20,32,33. Здесь мы оптимизировали очистку ядра ORF2p (остатки 238-1061) для получения миллиграммовых количеств более чем 99% чистого фермента (рис. 1c), который был мономерным (расширенные данные рис. 1a) и высокоактивным в отношении олиго(A) шаблонов (расширенные данные рис. 1b), что позволило провести структурный и кинетический анализы, а также анализ с разрешением а одно основание с различными субстратами и ингибиторами...
Discussion
Наши комплексные анализы раскрывают внутреннюю работу молекулярной машины, которая написала почти половину генома человека. Понимание структуры и функции L1 важно как для эволюции, так и, все чаще, для заболеваний человека. Накапливающиеся данные связывают активность L1 и ответ хозяина с распространенными патологиями, включая рак, старение, нейродегенерацию и аутоиммунитет2-7,26,27. Наши биохимические, структурные и эволюционные анализы показывают, что ORF2p содержит высокоактивную полимеразу, уникально приспособленную для паразитического цикла репликации, с сохраненными и новыми структурными особенностями, которые обеспечивают оптимальную ретротранспозицию на протяжении всей эволюции. В совокупности эти данные дают представление о двух ключевых механизмах, через которые L1 может вызывать заболевания: (1) ядерный инсерционный мутагенез и обусловленный им геномный хаос и (2) цитозольное считывание продуктов обратной транскрипции ORF2p.
Хотя активность ядерного L1 коррелирует с повреждениями ДНК и структурными геномными перестройками 2,41,42,52, механистическое понимание инсерции L1 остается неуловимым. Процесс вставки можно представить как две половинные реакции: синтез первой и второй нити. Синтез второй нити было сложно изучить, и было неясно, осуществляется ли он L1 или хозяином. Наши данные показывают, что фермент ORF2p компетентен выполнять все ферментативные шаги, необходимые для прайминга и осуществления синтеза как первой, так и второй нити: он эффективно синтезирует ДНК с короткими РНК или ДНК праймерами как на РНК, так и на ДНК шаблонах (рис. 3, расширенные данные рис. 4 и 5 и дополнительные рис. 3-5). Интерпретация наших результатов в контексте высококачественных биохимических данных, полученных за десятилетия изучения R2 LINEs у насекомых 21,24,36,48 , дает нам возможность обновить модель инсерции L1 (рис. 6). Этот механизм описывает каноническую инсерцию, которая намеренно упрощена и опускает многочисленные поддерживающие и репрессирующие белки хозяина, включая топоизомеразу TOP1, PARP1, белки, связывающие богатые пурином элементы, путь Fanconi (включая BRCA1) и p53 (ссылки 8,17,18,19). Кроме того, альтернативные пути, такие как катализируемый хозяином синтез второй нити, могут происходить в различных контекстах или после сбоя ORF2p, и хозяин может бороться со вставкой, например, расщепляя промежуточные продукты.
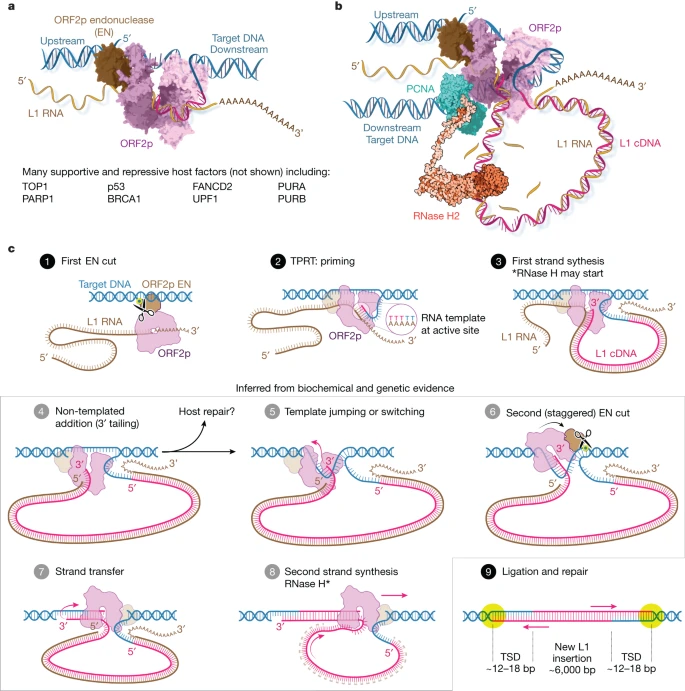 Fig. 6: Revised L1 insertion model.
a, ORF2p bound to target DNA as TPRT begins, drawn schematically with linear target DNA for clarity as in the models below. b, ORF2p in complex during first strand synthesis. It seems more likely that ORF2p bends the target DNA around the highly positively charged ‘back’ face of the polymerase (Extended Data Fig. 9); it can then pass through the PCNA ring clamp, which binds to the PIP box and recruits RNase H2 (ref. 29). c, Revised insertion model. Activities supporting steps 4, 5, 7 and 8 are demonstrated here. 1. ORF2p EN cuts target DNA, liberating a gDNA 3'-OH 2. TPRT: the T-rich gDNA primer is passed into the RT active site, where it base pairs with the poly(A) tail of the bound template, and the 3'-OH is extended. 3. First strand synthesis generates a large (6 kb) cDNA loop; RNase H2, recruited by ORF2p–PCNA, can begin. 4. NTA, in which extra bases are added to the 3' cDNA end beyond the 5' end of the RNA template, may occur. 5. Template jumping or switching to the exposed single-stranded gDNA may follow, potentially facilitated by microhomology from NTA nucleotides and the 5' cap. This would also release 5' phosphate-bound EN to ‘rock and roll’20,24,48 to carry out: 6. The second EN (staggered) cut, which liberates the 3' OH used to prime second strand synthesis; a stagger from the first cut of approximately 12–18 bp results in characteristic target site duplications (TSDs)20,21,24,44. 7. Strand transfer and priming of second strand synthesis. 8. Second strand synthesis using the 6 kb L1 cDNA as template. RNase H2 activity may also occur here. 9. Ligation and end repair, resulting in a completed approximately 6 kb insertion flanked by TSDs. The second EN cleavage may sometimes occur in the absence of a template jump. b, © 2023 JHUAAM. Illustration: Jennifer E. Fairman.
Наши данные также проливают свет на другие области канонического цикла репликации L1. Считается, что связывание цис-РНК ORF2p происходит на рибосоме53,54. Вновь транслированный апо ORF2p нестабилен до связывания РНК и принимает конформацию "большой палец вверх", способную плотно связывать РНК; мы предполагаем, что первоначальное связывание РНК, вероятно, происходит одновременно с трансляцией, потенциально еще до трансляции CTD. Связывание PCNA, которое необходимо для ретротранспозиции17 и рекрутирует РНКазу H2 для обеспечения расщепления второй нити29, по-видимому, не блокируется ни в одном из идентифицированных состояний; это, наряду с зависимостью от EN и RT17,18, указывает на то, что PCNA может быть рекрутирована в ORF2p в результате развивающегося геномного повреждения. Большинство новых LINE-вставок сильно усечены по 5'1; часто они состоят всего из нескольких сотен пар оснований, но причины этого не вполне понятны. ORF2p эффективна и высокопроцессивна, что согласуется с предыдущими наблюдениями16,32, это подтверждает идею о том, что расщепление хозяином РНК L1 или промежуточных продуктов с большей вероятностью вызывает усечение 5', чем неэффективность полимеразы55. Уровень ядерного ORF1p ограничен17,18, и связанные шапероны ORF1p будут вытеснены из L1 РНК во время RT, потенциально оставляя незащищенным большой одноцепочечный промежуточный цикл кДНК (шаги 3-7, рис. 6). Учитывая его гомологию с большей частью генома, он может представлять собой как уникальную уязвимость, так и потенциальный очаг транслокаций41,42,52.
Известно, что цитозольные двуцепочечные нуклеиновые кислоты, вирусная мимикрия и обусловленная ими интерфероновая сигнализация способствуют развитию патологии в различных контекстах, и было показано, что NRTI ограничивают выработку интерферона и этих нуклеиновых кислот6,7, однако их происхождение остается спорным. Во-первых, наши данные показывают, что ORF2p может использовать РНК-праймеры и короткие РНК-шпильки для запуска реакций RT; Alu-подобная последовательность легко растягивается, а уридилирование РНК L156 может также превратить ее в аналогичный субстрат. РНК-прайминг ORF2p RT в цитоплазме позволяет объяснить происхождение этих нуклеиновых кислот с достаточной степенью вероятности. Мы также показали, что праймеры ДНК длиной 5 нт могут праймировать L1; возможно, что допускаются и более короткие праймеры16. Во-вторых, мы показали, что L1 может напрямую синтезировать гибриды РНК:ДНК в цитозоле; это RT-зависимо, но EN-независимо, что исключает ядерное происхождение этой системы. В-третьих, мы показали, что синтезированные L1 кДНК активируют cGAS/STING, что приводит к выработке интерферона. Наши наблюдения также демонстрируют потенциально критическую роль L1 и его RT-продуктов в вирусной мимикрии57,58, что следует из эволюции генома и рака59,60. Более того, наши данные о надежных ингибиторах создают основу для оценки участия L1 в этих фенотипах и для воздействия на него в будущем. Таким образом, структурное раскрытие ORF2p облегчит рациональный дизайн новых терапевтических средств и заложит основу для будущих исследований, необходимых для изучения и углубления понимания механизма вставки L1, его эволюции и роли в развитии заболеваний.
|