|
|
|
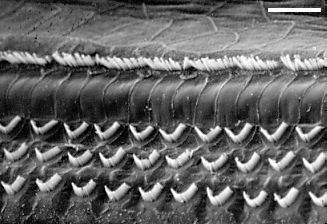
| У человека дифференцировка Кортиевого органа начинается на 9 неделе беременности (wg). Показаны основные стадии развития (refs. a4, c3, c4). Время соответствия между человеком и разными экспtриментальными животными приведено в таблице I . |
| |||||||||
| ||||||||
| |||||||||
|
|
| |
 |
 |
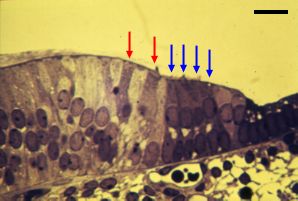
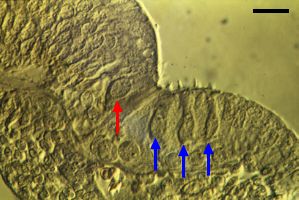
| Избыточные волосковые клетки часты и временами сталкиваются во время развития Кортиева органа человека между стадией 3 и 4 (см. ref. c4). Как здесь(14 wg, слева; 20 wg, справа) xfcnj dblys 2 ряда IHCs (красные стрелки) и 4 или 5 рядов OHCs (голубые стрелки). bar: 20 µm | |
| |||||||
| Эта таблица позволят сравнить время стадии 2 ( начало морфологической дифференцировки волосковых клеток),
стадии 5 (начало функционирования улитки), и стадии 6 (конец созревания) у разщных млекопитающих(ref.c1#c1). wg = неделя беременности(человек) ; ed = день эмбриогенеза ; pnd = день постнатального развития | |||
|
|
|
|
|
| human rat, mouse cat guinea-pig gerbil |
10 wg 16-17 ed ? ed 34 ed ? ed |
18 wg 8-10 pnd 3 pnd 54 ed 12 pnd |
30 wg 16-20 pnd 20 pnd 6 pnd 20 pnd |
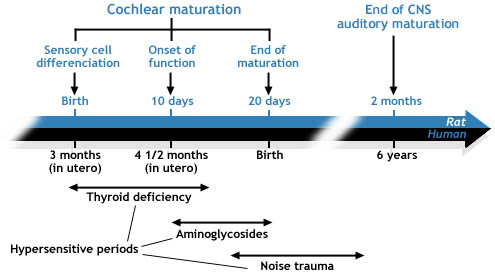 |
| На таблице представлены стадии развития 2, 5, и 6 (см. выше) созревания улитки и отмечено более позднее развитие слуховой части головного мозга. Действительно, для собственного созревания головной мозг нуждается в том, чтобы улитка полностью созрела и была функциональной. Показаны периоды гиперчувствительности к 3-м основным повреждающим развитие улитки факторам. Во время этих периодов улитка наиболее чувствительна, по сравнению со взрослой (ref.a2, b1). |
|
Reviews a2. a name=a2>PUJOL R., UZIEL A. Auditory development : peripheral aspects. In: Meisami E, Timiras PS. Handbook of human growth and developmental biology, vol I part B. Boca Raton, Florida: CRC Press, pp. 109-130, 1988. a3. LIM D.J., RUEDA J. Structural development of the cochlea. In: Romand (ed) Development of auditory and vestibular systems 2. Oxford: Elsevier pp. 33-58, 1992. a4. a name=a4>PUJOL R., LAVIGNE-REBILLARD M., LENOIR M. Development of sensory and neural structures in the mammalian cochlea. In: Rubel E.W., Popper, A.N., Fay R.R. eds. Development of the auditory system, Springer Handbook of Auditory Research, vol 9, pp 146-192, 1997.
b2. ROMAND R. ed. Development of Auditory and Vestibular systems. Academic Press, New-York, 1983. b3. ROMAND R. ed. Development of Auditory and Vestibular systems 2. Elsevier, Oxford, 1992. b4. RUBEL E.W., POPPER A.N., FAY R.R. eds. Development of the auditory system, Springer Handbook of Auditory Research vol. 9, Springer, New York, 1997 c1.a name=c1> PUJOL R., and HILDING D. Anatomy and physiology of the onset of auditory function. Acta Otolaryngol (Stockh) 76:1-11, 1973 c2. LENOIR, M., SHNERSON, A. & PUJOL, R. (1980) Cochlear receptor development in the rat with emphasis on synaptogenesis. Anat. Embryol. 160, 253-262 c3.PUJOL R., LAVIGNE-REBILLARD M. Early stages of innervation and sensory cell differentiation in the human fetal organ of Corti. Acta Otolaryngol (Stockh) Suppl 423:43-50, 1985 c4. a name=c4>LAVIGNE-REBILLARD M, PUJOL R. Development of the auditory hair cell surface in human fetuses. A scanning electron microscopy study. Anat Embryol 174:369-377, 1986 c5. LENOIR M., PUEL J.L., PUJOL R. Stereocilia and tectorial membrane development in the rat cochlea. A SEM study. Anat Embryol 175:477-487, 1987 Сайт создан в системе uCoz
|

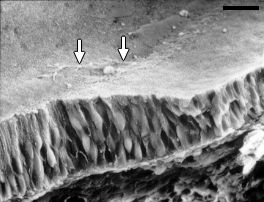

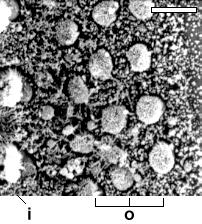
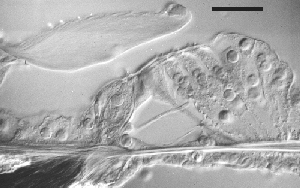
 Nomarski срез улитки 11 wg плода .
bar: 20
µm
Nomarski срез улитки 11 wg плода .
bar: 20
µm

 У всех изученных млекопитающих эта стадия соответствует
У всех изученных млекопитающих эта стадия соответствует