Текториальная мембрана (TM) является спиральной лентой из внеклеточного матрикса, которая располагается над Кортиевым органом. Она прикреплена своим медиальным краем к верхушке спирального лимба (spiral limbus), вытягивается поперек внутренней бороздки, и располагается поверх внутренних и наружных воловсковых клеток. Ряд структурных специализаций можно различить на текториальной месбране. На верхней поверхности имеется , аностомозирующая сеть толстых тяжей (cables). На латеральном крае имеется , который может быть прикреплен посредством маргинальной сети к верхушкам последнего ряда Дейтеровских клеток. На нижней поверхности имеется
, выступающий гребень, который идет продольно вдоль текториальной мембраны и предназначен для контакта и управления пучками внутренних волосковых клеток . Самые высокие в нижнюю поверхность текториальной мембраны, эта область известна как Кimura's мембрана.
ТМ содержит радиально расположенные пучки коллагена, встроенные в фибриллярный матрикс.
Разрез типичного базального витка улитки млекопитающих. Диаметр наружной волосковой клетки примерно 7 µm. Пустые пространства в основаниях наружных волосковых клеток заняты эфферентными окончаниями, которые не видны на рис. 1 - базилярная мембрана, 2 - Hensen клетки, 3 - наружные фалангеальные клетки Deiters, 4 - нервные окончания, 5 - наружные волосковые клетки, 6 - наружные спиральные волокна, 7 - наружные pillar клетки, 8 - внутренний туннель, 9 - внутриенние pillar клетки, 10 - внутренние фалангиальные клетки, 11 - пограничные клетки, 12 - текториальная мембрана, 13 - клетка спирального ганглия типа I, 14 - клетка спираьного ганглия типа II, 15 - костная спиральная ламина, 16 - спиральный кровеносный сосуд, 17 - веретенообразные клетки, 18 - аксоны клетки спирального ганглия (волокна слухового нерва), 19 - радиальные волокна.
SEM разлом ТМ
Эта сканированная электронная микрофотография разлома через Кортиев орган показывает покрывающую сеть на верхней поверхности ТМ и маргинальный диск (красные стрелки), который идет вдоль латерального края. На границе разлома видны OHCs и фалангеальные отростки Дейтеровских клеток . bar: 20
µm
Маргинальная сеть присоединяет ТМ к наружному ряду Дейтеровских клеток ( здесь невидны): см. .
TEM покровная сеть тяжей (cable) (слева) и маргинальный диск (справа)
Трансмиссионная электронная микрофотография среза через один из кабелей покровной сети (слева) и через маргинальный диск (справа). Тонкие фибриллы ТМ матрикса компактны и сложены тесно друг к другу благодаря этому образуют электронплотную структуру. bars:0.2 µm (left) ; 1 µm (right)
Гензеновская полоска (Hensen's
stripe)
Трансмиссионная электронная микрофотография среза через Гензеновскую полоску. Тонкие плотно упакованные фибриллы формируют клиновидный гребень, который по всей своей длине контактирует с кончиками пучков стереоцилий внутренних волосковых клеток.
bar: 1 µm
Отпечатки стереоцилий
Сканирующая электронная микрофотография показывает пучки наружных волосковых клеток в Кортиевом органе и нижнюю поверхность текториальной мембраны этой области. Текториальная мембрана отсоединена от пучков наружных волосковых клеток во время приготовления препарата. На этой нижней поврехности ТМ видные отпечатки пучков стереоцилий волосковых клеток. bar: 10 µm
см. также между ОНС и ТМ
TEM срез через средину тела текториальной мембраны .
Видны радиальные пучки прямых , 20 nm в диаметре коллагеновых фибрил, включенных в фибриллярный матрикс, образуемый двумя типами тонких диаметром (7-9 nm) фибрил светло и темно окрашенного типа() . Эти светло и темно окрашенные тонкие в диаметре фибрилы располагаются альтернативно и связаны друг с другом эшелонироваными поперечными мостиками для образования того, что м.б. описано как полосатый слой (striated sheet) матрикса
().
bar: 200 nm
TEM срезов через средину тела ТМ, которая была обработана буффером ( слева) или буффером. содержащим бактериалрную коллагеназу (справа) перед фиксацией. Фибрилы в 20 nm в диаметре чувтвительны к колагеназе, а striated sheet матрикса нет. Гель электрофорез показывает ТМ состоит из коллагенов и нечувствительных к колагеназе гликопротеинов. bar: 0.2
µm
SDS гель белков текториальной мембраны
(кликни на миниатюре, чтобы увидеть в реальном размере картину и подпись).
alpha-tectorins cDNAs
кДНК текторинов (tectorins) клонирована (). Имеется большой связанныый дисульфидными мостиками комплекс, происходящий из одиночного большого (239,034 Da) полипептида, состоящего из 3-х отдельных модулей. Имеется NH2 область сходная с G0 доменом энтактина (entactin (ent, розовый)), центральная область с двумя частичными и тремя полными von Willebrand type D повторами(0-4, красный), который сходен с мембранным белком спермиев зонадгезином (zonadhesin), и С-терминальная область, которая формирует домен zona pellucida (zp, зеленый). Beta tectorin небольшой полипептид (36,074), содержащий одиночный домен zona pellucida domain (zp, зеленый). Два белка могут взаимодействовать homophically через посредство своих zona pellucida доменов и формировать гомомерные филаменты.
Модель образования гомомерных филамент и взаимодействий
Zona pellucida домен является элементом общим раду различных белков, каждый из которых или может или формирует филаментозные в основе внеклеточные структуры . Предполагается, что alpha и beta-tectorin образуют гомомерные филаменты посредством своих zona pellucida доменов (zp, зеленые). Центральный модуль alpha tectorin, обладает идентичностью с мембрнным белком спермиев zonadhesin, который взаимодействует с zona pellucida ооцитов. Гомомерные alpha и beta tectorin филаменты могут следовательно взаимодействовать посредством zonadhesin-подобного домена (vwf, красный) alpha текторина, чтобы образовать исчерченный слой (striated sheet) матрикса (см.
ниже)
TEM , показывающий детали striated
sheet matrix. Обратите внимание на филаменты светлого и темного типа и стойкие поперечные мостики, связывающие оба .
bar: 50
nm
Reviews
a1. Lim DJ (1986) Functional structure of the organ
of Corti: a review.
a2.
...and also:
c1. Richardson GP, Russell IJ, Duance VC, Bailey AJ
(1987) Polypeptide composition of the mammalian tectorial membrane.
c2.
Thalmann I, Thallinger G, Crouch EC, Comegys TH, Barrett N, Thalmann R
(1987) Composition and supramolecular organisation of the tectorial
membrane.
c3. Hasko JA, Richardson GP (1988) The
ultrastructural organisation and properties of the mouse tectorial
membrane matrix.
c4. Slepecky NB, Cefaratti LK, Yoo TJ (1992) Type II
and type IX collagen form heterotypic fibers in the tectorial membrane of
the inner ear.
c5. Killick R, Legan PK, Malenczak C, Richardson GP
(1995) Molecular cloning of chick b-tectorin, an extracellular matrix
molecule of the inner ear.
c6. Killick R, Richardson GP (1997) Antibodies to
the sulphated, high molecular mass mouse tectorin stain hair bundles and
the olfactory mucus layer.
c7. Legan PK, Rau A, Keen JN, Richardson GP (1997) The mouse
tectorins: Modular matrix molecules of the inner ear homologous to
components of the sperm-egg adhesion system.


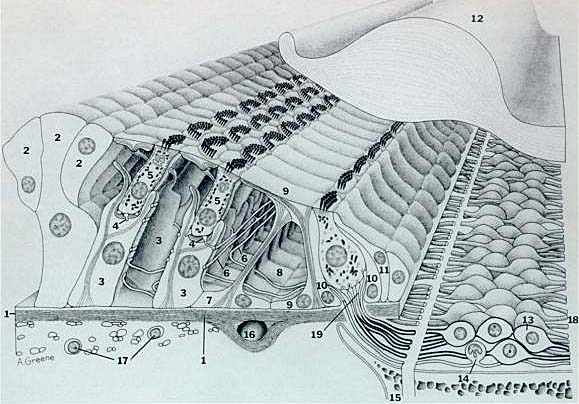 Разрез типичного базального витка улитки млекопитающих. Диаметр наружной волосковой клетки примерно 7 µm. Пустые пространства в основаниях наружных волосковых клеток заняты эфферентными окончаниями, которые не видны на рис. 1 - базилярная мембрана, 2 - Hensen клетки, 3 - наружные фалангеальные клетки Deiters, 4 - нервные окончания, 5 - наружные волосковые клетки, 6 - наружные спиральные волокна, 7 - наружные pillar клетки, 8 - внутренний туннель, 9 - внутриенние pillar клетки, 10 - внутренние фалангиальные клетки, 11 - пограничные клетки, 12 - текториальная мембрана, 13 - клетка спирального ганглия типа I, 14 - клетка спираьного ганглия типа II, 15 - костная спиральная ламина, 16 - спиральный кровеносный сосуд, 17 - веретенообразные клетки, 18 - аксоны клетки спирального ганглия (волокна слухового нерва), 19 - радиальные волокна.
Разрез типичного базального витка улитки млекопитающих. Диаметр наружной волосковой клетки примерно 7 µm. Пустые пространства в основаниях наружных волосковых клеток заняты эфферентными окончаниями, которые не видны на рис. 1 - базилярная мембрана, 2 - Hensen клетки, 3 - наружные фалангеальные клетки Deiters, 4 - нервные окончания, 5 - наружные волосковые клетки, 6 - наружные спиральные волокна, 7 - наружные pillar клетки, 8 - внутренний туннель, 9 - внутриенние pillar клетки, 10 - внутренние фалангиальные клетки, 11 - пограничные клетки, 12 - текториальная мембрана, 13 - клетка спирального ганглия типа I, 14 - клетка спираьного ганглия типа II, 15 - костная спиральная ламина, 16 - спиральный кровеносный сосуд, 17 - веретенообразные клетки, 18 - аксоны клетки спирального ганглия (волокна слухового нерва), 19 - радиальные волокна.










