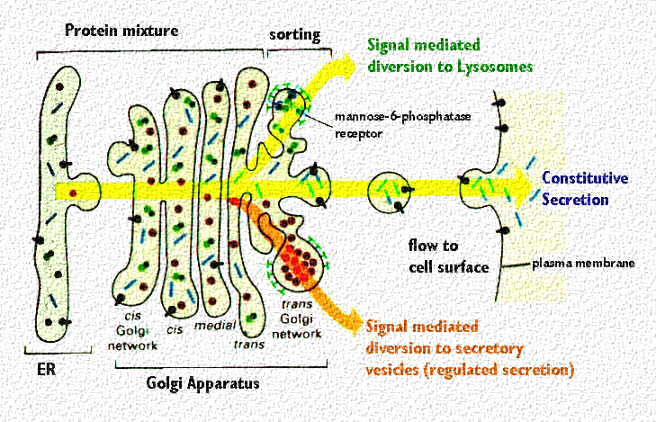
На схеме показана сортировка белков, предназначенных для секреции, в вакуоли в trans комплексе. Обратите внимание, что лизосомальные энзимы сортируются в другом компартменте, это рецепторы mannose 6 phosphate.
ORIGINAL RESEARCH PAPER Marsh, B. J. et al. Organellar relationships in the Golgi region of the pancreatic beta cell line, HIT-T15, visualized by high resolution electron tomography. Proc. Natl Acad. Sci. USA 98, 2399-2406 (2001). | | | |
is described by its creators as "a general computational framework for modelling cell biological processes". Access to this software is available over the internet through a JAVA-based interface
Получена трехмерная реконструкция области вокруг аппарата Гольджи панкреатической β-клетки (HIT-T15)
Organelles in the Golgi region. C1 (cis-Golgi cisterna), light blue; C5, dark blue; C7, red; ER, yellow; microtubules and mitochondria, green; dense core vesicles, blue; clathrin-negative vesicles, white; clathrin-positive vesicles and compartments, red; clathrin-negative compartments, purple; free ribosomes, orange.
Не обнаружено ничего необычного в самом аппарате Гольджи, который здесь состоит из семи цистерн (C1 - C7, идущих от cis к trans Golgi). Интересным оказалось наблюдение, что endoplasmic reticulum (ER), по-видимому, пронзает массу Гольджи через упорядоченно расположенные отвестия в цистернах. Имеется тесный контакт между ER и цистернами C5, C6 и C7, которые м.б. точками прямого обмена между этими органеллами, минуя везикулярный транспорт.
Микротрубочки, по-видимому, следуют близко к мембране C1 цистерны аппарата Гольджи и эндосомных компартментов. Однако, ER, по-видимому, закреплен вдоль микротрубочек лишь в нескольких точках.
Считалось, что область Гольджи сильно переполнена, однако, Marsh и др. подсчитали, что лишь 34% объема занято органеллами, остальная часть занята цитоплазматическим матриксом.
Mobile
Factories: Golgi Dynamics in Plant Cells - reviews recent
advances in understanding the machinery dynamics of the Golgi
apparatus in plant cells. From Trends in Plant Science, 2001,
6:4:160-167. Full text available from
BioMedNet.
Organization
of the Golgi Apparatus - emphasizes cross-species comparisons.
From Current Opinion in Cell Biology, 2000, 12:450-456. Full
text available from BioMedNet.
Transport
between ER and Golgi - focuses on the mechanisms and regulation
of transport. From Current Opinion in Cell Biology, 2000,
12:445-449. Full text available from
BioMedNet.
Regulators
and Effectors of the ARF GTPases - discusses how ARFs regulate
membrane dynamics within the cell. From Current Opinion in Cell
Biology, 2000, 12:475-482. Full text available from
BioMedNet.
- offers fascinating movies of intracellular
movement made possible by the use of the green fluorescent protein.
From the July 1998 issue of Molecular Biology of the
Cell.
- provides background
information and diagrams.
Benjamin S. Glick CAN THE GOLGI FORM DE NOVO? Nature Reviews Molecular Cell Biology 3, 615 -619 (2002)
Недавние исследования показали, что аппарат Гольджи м. формироваться de novo с помощью самоорганизации.
(Рис.2.) | A hypothetical mechanism for de novo Golgi formation.
Golgi biogenesis in yeast
The Golgi Apparatus Going Round in Circles? Francis A. Barr HMS Beagle,TRENDS PREVIEW 2,Posted February 1, 2002 · Issue 119
Считается, что транспорт белков из endoplasmic reticulum (ER) и через Golgi aппарат обеспечивается с помощью небольших
пузырьков переносчиков, отшнуровывающихся от одного
компартмента и сливающихся со следующим []. Это мнение поставлено под сомнение
при исследовании секреции проколлагена, молекул очень больших по сравнению с классическими пузырьками носителями 60-70 nm
диаметром, которые проходят антероградно через Golgi стек (stack), не покидая цистерн []. Модель cisternal maturation используется для объяснения этих данных (Рис.1.) | . Согласно этой модели, цистерны Golgi возникают в результате слияния везикулярных или тубулярных промежуточных структур из ER и затем созревают из цис- в транс-форму путем обмена энзимами с малыми везикулярными носителями, движущимися ретроградным способом. Mironov и др. расширили свои наблюения с проколлагеном, включив значительно меньший трансмембранный гликопротеин vesicular stomatitis virus (VSV-G) []. Они установили, что никакие молекулы груза не вступают в транспортные пузырьки, чтобы пройти
через массу аппарата Гольджи и что
этот груз транспортируется с той же
самой скоростью, которая характерна
для созревающих структур цистерн.
Биохимические и морфологические
доказательства подтверждают и др.
аспекты модели созревания цистерн.
Установлено, что находящиеся в
Гольджи белки рассортировываются
по recycling пузырькам на разных
уровнях Golgi stack []. Исследования лаб. Nilsson описывают
очистку разных классов COP I
пузырьков, содержащих или цис-
или medial-Golgi маркеры, но
содержащих мало антероградного груза []. Эти пузырьки отбирают груз путем
купирования рекрутируемых COP I
комплексов, чтобы гидролизовать
ГТФ с помощью малой GTPase ARF1. Этот
механизм, по-видимому, связан с
вовлечением ARF1 в прямую
модуляцию активности GTPase-activating protein (GAP)
с помощью, по крайней мере трансмембранных грузовых молекул. Цитоплазматические домены этих
белков м. ингибировать активность
ARF-GAP, позволяя тем самым накапливаться активным ARF1 на мембранах, и таким образом способствуя рекрутированию COP I и
образованию пузырьков [].
Итак, некоторые, если не все, антероградные грузы
транспортируются в созревающих
цистернах, тогда как энзимы присутствуют в COP
I пузырьках, подтверждая модель
созревания для Гольджи транспорта.
При этом возникает вопрос поддержания сложившейся
цистерновой структуры аппарата
Гольджи. Модель maturation предполагает,
что структурные белковые комплексы
, часто обозначаемые как "Golgi matrix,"
должны быть высоко динамичными,рециклировать по мере созревания цистерн, тогда как раньше они рассматривались как довольно
статичные компонеты длительно
живущих цистерновых структур.
The Golgi Matrix: Is This Dynamic or Static?
Предполагаемый Golgi matrix был первоначально описан биохимически
и морфологически как белок-содержащая
стуркура, выделяемая из мембран Гольджи, которая поддерживает форму цистерн Гольджи и способна специфически связываться с энзимами Гольджи (Рис.1.) | []. Очищенные мембраны Golgi сохраняют свою характерную составленную из цистерн структуру, поэтому было предположено, что белки матрикса
Гольджи формируют экзоскелет,
необходимый для поддержания этой
уникальной организации. Многие
компоненты этого матрикса Гольджи являются
супер-скурченными (coiled-coil) белками,
лучше всех из них охарактеризованы cis-Golgi
белки GM130 и взаимодействующий с ним
партнер p115 []. GM130 направляется в мембраны Гольджи
благодаря взаимодействию с RASP65, белком, идентифицированном в in vitro функциональном тесте как фактор,
необходимый для укладывания
цистерн Гольджи []. Интересно, что GRASP65 является myristoylated
белком и что многие др. белки с подобной модификацией м. делать выбор между мембран-связанной и цитозольной формой в ответ на
триггер, такой как ГТФ связывание []. Исследования динамики GFP-нагруженной
формы GRASP65 in vivo показали, что она, по-видимому, быстро рециклирует на и вне мембран
Гольджи []. Остается или нет она при этом
связанной с GM130 в этих экспреиментах
выяснить не удалось, но предварительные биохим.
исследования показали, что GM130 и GRASP65
взаимодействую.т с высоким сродством, это делает очень
вероятным предположение, что комплекс рециклируется как
целое []. Это исследование также показало,
что имеется цитозольный пул GRASP65.
Было также установлено, что вновь
синтезируемые эндогенные GRASP65 и GM130
быстро и непосредственно связываются и с Golgi, но в этих экспреименах не получено доказательств существования
цитозольного пула для этих двух белков []. Одна возможность, что GRASP65-GM130 комплекс активно
рециклируется из мембран Гольджи
в цитозоль, где имеется лишь очень
небольшой пул в любой момент
времени. Наконец, необходимо
помнить, что обмен ассоциированных
с мембранами субъединиц матрикса
Гольджи с растворимым пулом, сам по
себе не доказывает, что цистерны
меняют свою идентичность.
Does the Golgi Matrix Exist?
Warren с сотр. нашли, что существуют
Гольджи-подобные структуры
независимо от транспорта белков из
эндоплазматического ретикулема и
представили доказательства
сущестования ригидного Гольджи-матрикса []. В их экспреиментах эти матричные
структуры , напоминающие Гольджи,
видны при световой микроскопии, но
не видны при ЭМ, у них отсутствует
четкая цистерновая организация []. Очевидно, что не только Гольджи-матрикс
необходим для образования аппарата
Гольджи, хотя эти данные и подтверждают идею, что матрикс
Гольджи, структура независимая от ER.
Используя микроинъекции или избыточную экспрессию доминантно-негативной мутации Sar1 для блокирования выхода
из ER с помощью COP-II везикулярного
транспортного пути [], было показано, что Golgi белки
перераспределяются в ER, а аппарат Golgi
исчезает. Кинетика релокализации
была различной для Гольджи энзимов
по сравнению с Golgi matrix белками;
последние перераспределялись более
медленно и этот процесс нуждался в
высоких концентрациях dominant-negative Sar1 []. Эти эксперименты ставят под вопрос
независимость матрикса от ER. Однако,
предостережением этому является то,
что это не доказывет, что матричные
белки Гольджи рециклируют обратно в
ER в нормальных условиях. Кроме того
необходимо много часов для
матричных Гольджи белков, чтобы
перераспределиться в ER , а в
короткое время не обнаруживается
эффекта на матричных белках GM130 и GRASP65 []. Однако, перераспределение др.
матричных белков Гольджи протекало
минуты в сходных условиях [].
GM130, как известно, взаимодействует как с Golgi Rab белками, так и с фактором cвязывания пузырьков p115 и играет
роль в формировании Golgi stacks в
бесклеточных условиях []. Новый Golgiматричный белок, golgin-45,
является связывающим партнером
для GRASP55 в medial-Golgi []. Хотя функция golgin-45 не известна, он
связывается с малой GTPase rab2, участвуя
в некоторых аспектах привязывания пузырьков к medial-Golgi. Устранение golgin-45 с помощью RNA
interference ведет к коллапсу энзимов
Гольджи обратно в ER, а др. Golgi
матричные белки при этом перераспределяются в небольшие
точечные структуры и цитоплазму.
Не удивительно, что это также
вызывает блокирование транспорта
белков из-за отсутствия функционального аппарата Гольджи.
Итак, хотя Golgi матричные белки, по-видимому,
важны для поддержания структуры Гольджи для предопределния идентичногости Гольджи, существование упорядоченной
структуры , обозначаемой matrix , все еще неясно. Вряд ли этот статичный матрикс, необходимо принимать во внимание
альтернативную модель, согласно
которой эти белки существуют в виде
небольших landmark комплексов,
специфичных для разных цистерн
Гольджи, таких как GRASP65-GM130-p115
комплекс на cis-Golgi, и GRASP55-golgin-45 на medial-Golgi, чтобы обеспечить идентификацию компартментов (Рис.2.) | .
Concluding Remarks
Все еще имеются нерешенные вопросы рассматриваемой модели.
Известно, что Гольджи энзимы рециклируют посредством ER и что весь перемещаемый вперед груз, по-видимому,
включает энзимы, двига.щиеся с
той же самой скоростью через
аппарат Гольджи. Если это так, то cis-Golgi
должен содержать не только cis-энзимы,
но и значиетльное количество
рециклированных medial- и trans-энзимов
на их пути назад в их соответствующее местоположение.
Как затем осуществляется их
дискретное распределение по разным
слоям Гольджи ? Возможно, что существует механизм
последовательного рециклинга
энзимов, который транспортирует
энзимы из предшествующих цистерн в
пузырьки скорее, чем непосредственно позволяет
возвращаться назад в ER. Если эти
пузырьки не диффундируют свободно,
а привязаны к соседним компартментам предназначения с
момента их образования, то это м. объяснить такой механизм.
Еще один вопрос, связанный с предполагаемыми Golgi матричными
белками, как они специфически попадают в цель и сохраняют свою локализацию в разных цистернах
внутри агрегата. Возможно, что
некоторые матричные белки рециклируют в цитоплазме, но др. являются интегральными мембранными белками и д., следовательно,
рециклироваться в пузырьках.