A bipotential precursor population for pancreas and liver within the embryonic endodermGail Deutsch, Joonil Jung, Mighua Zheng, Jose Lora and Kenneth S. Zaret
Development V.128. P. 871-81. 2001 (Перевод В.К. Викуловой) |
Поджелудочная железа выделяется независимо от дорзального и вентрального доменов эмбриональной кишечной энтодермы. В опытах по инактивации генов мыши были идентифицированы факторы, необходимые для развития дорзальной части поджелудочной железы, но факторы, инициирующие развитие вентральной части, остались невыявленными . В этой работе исследуется предположение, что выделение вентральной части поджелудочной железы взаимосвязано с выделением печени. Обнаружено, что спецификация печени и вентральной части поджелудочной железы происходит одновременной в одном и том же общем домене клеток. В опытах по эксплантации эмбриональной ткани был обнаружен дефолт(невозможность продолжения) пути развития энтодермы вентральной части передней кишки, т.е. неспособность к активации генной программы поджелудочной железы. Сигналы FGF от кардиальной мезодермы стимулируют эту энтодерму экспрессировать гены печени вместо генов поджелудочной железы. Не найдено никаких доказательств о том, что на выбор типа клеток поджелудочной железы или печени влияет избирательный рост или жизнеспособность клеток. Кардиальная мезодерма или сигнал FGF индуцируют локальную экспрессию sоnic hedgehog(Shh), который, в свою очередь, подавляет развитие поджелудочной железы, но не печени. Популяция бипотенциальных клеток-предшественников поджелудочной железы и печени в эмбриональном развитии и их выбор пути развития с помощью FGF имеет черты, которые являются, по-видимому, повторением особенностей поджелудочной железы, возникавших в ходе эволюции.
Механизмы передачи сигналов, контролирующих спецификацию органов-производных энтодермы таких, как поджелудочная и щитовидная железы, печень, легкие и кишечник , начали исследовать недавно. Много известно о том, как органы растут, проходят дифференциацию и морфогенез (Hogan,1999), но как разные домены энтодермы кишки дают начало этим органам на начальном этапе известно мало(Wells and Melton, 1999; Gannon and Wright, 1999; Zaret, 2000). В этой работе исследована способность общих клеток-предшественников поджелудочной железы и печени к развитию и способа выбора ими одного определенного клеточного типа.
Поджелудочная железа происходит от особых эмбриональных выростов дорзальной и вентральной областей энтодермы передней кишки, выростов, дающих начало эндокринным и экзокринным клеткам(Spooner et al., 1970; Slack, 1995). Печень же развивается исключительно из вентральной энтодермы передней кишки(Zaret, 1996), расположенной рядом с местом, где выделяется вентральная часть поджелудочной железы. Многие доказательства свидетельствуют о существовании различий в процессах спецификации дорзальных и вентральных зачатков поджелудочной железы. У цыпленка сигналы от покрывающей сверху нотохорды активизируют развитие дорзальной части поджелудочной железы путем подавления энтодермальной экспрессии sonic hedgehog (Kim et al., 1997; Hebrok et al., 1998; Apelqvist et al., 1997; Kim and Melton, 1998); но нотохорда не влияет на экспрессию генов в вентральном зачатке поджелудочной железы (Kim et al., 1997; Hebrok et al., 1998). Опыты по изучению генной активации у мышей показали, что развитие дорзального зачатка поджелудочной железы зависит от транскрипционных факторов Isl1 и Hlxb9; вентральный зачаток поджелудочной железы все еще продолжает развиваться, когда эти гены инактивированы (Ahlgren et al., 1997; Нarrison et al., 1999; Li et al., 1999). Параметры спецификации вентральной части поджелудочной железы неизвестны.
Опыты по эксплантации ткани показали, что спецификация вентральной части поджелудочной железы происходит на стадии 7-9 сомитов(Wessеlls and Cohen, 1967), тогда как морфогенез вентрального зачатка поджелудочной железы не определяется до стадии 20 сомитов( Spooner et al., 1970). Об экспрессии известных ранее панкреатических маркеров Hlxb9 и гомеобоксного белка Pdx1 (Jonsson et al., 1994; Guz et al., 1995; Offield., 1996) в вентролатеральной части передней кишки на стадии 8 сомитов (Li et al., 1999) уже сообщалось. Эти транскрипционные факторы, вероятно, отвечают на первичные сигналы для спецификации поджелудочной железы и обозначают стволовые клетки поджелудочной железы до начала морфогенеза. Pdx1 необходим для морфогенеза и дифференциации эпителия поджелудочной железы после инициации экспрессии глюкагона и инсулина на стадии почки (Offield et al., 1996; Algren et al., 1996). Несмотря на то, что домен инициальной экспрессии Pdx1 включает часть будущего желчного протока и дуоденум (Offield et al., 1996), Pdx1-положительные клетки дают начало Isl1 и ngn3-положительным клеткам как предшественникам эндокринной линии (Ahlgren et al.,1997; Gradwohl et al., 2000) и р48-положительным клеткам как предшественникам экзокринной линии (Krapp et al., 1998).
В работах с использованием клеточного маркирования показано, что эндокринная и экзокринная линии клеток возникают из энтодермы (Percival and Slack, 1998). Также при изучении химерных мышей установлено, что островки эндокринных клеток по происхождению поликлональны (Deltour et al., 1991). Основываясь на этих фактах, можно предположить, что не связанные непосредственным клональным родством энтодермальные клетки, образуют популяцию стволовых клеток поджелудочной железы. Резуьтаты исследования роли сигналов от мезенхимы в развитии эндокринной и экзокринной линий с использованием эксплантации и трансплантации кусочков ткани зачатка поджелудочной железы , выделенных позже периода спецификации ( 15-25 сомитов и более; см. Wessels and Cohen, 1967; Rutter et al., 1978; Sanvito et al., 1994; Gittes et al., 1996; Miralles et al., 1998; Miralles et al., 1999), оставляют открытым вопрос о роли мезодермы в контроле формирования пакреатических стволовых клеток.
Временная и физическая близость локализации печени и вентральной части поджелудочной железы приводит нас к выяснению, каким образом спецификация этих двух тканей может быть связана. На стадии образования вентральной части передней кишки кардиальная мезодерма располагается рядом с энтодермой, и анализ результатов эксплантации и трансплантации ткани показал, что для развития вентральной энтодермы в направлении печени необходима близость к кардиальной мезодерме (LeDouarin, 1964; LeDouarin, 1975; Gualdi et al.,1996). У эмбрионов мыши взаимодействия между этими тканями происходят на стадии 7-8 сомитов, в период времени, когда специфические для печени гены активируются ( Cascio and Zaret, 1991; Shiojiori et al., 1991; Gualdi et al., 1996). При анализе энтодермальных эксплантатов выяснено, что для индукции гепатоцитов в энтодерме вентральной части передней кишки необходимо и достаточно избытка ростового фактора фибробластов (FGF) клеток кардиальной мезодермы( Jung et al., 1999) , являющегося ее сигналом( Jung et al., 1999).
В настоящей работе исследовано, как клетки поджелудочной железы и печени выделяются из вентральной части передней кишки. Результаты показывают, что выбор клеточного типа контролируют сигналы от кардиальной мезодермы. Прежде опубликованные наблюдения об изменении дифференциации клеток поджелудочных желез у взрослых иллюстрируют повторение выбора клеточного типа в развитии.
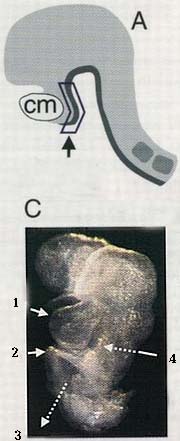
Рис. 1. (А) Вид с боковой стороны на эмбрион мыши на стадии 4-6 сомитов. Очерченный район соответствует анализируемой области; cm - кардиальная мезодерма. (С) Передняя половина 8-сомитного эмбриона. Обозначения: 1 - кардиальная мезодерма; 2 - энтодермальная губа; 3 - средняя кишка; 4 - пунктирная стрелка показывает, что энтодерма передней кишки находтся позади клеток кардильной мезодермы.
ОБСУЖДЕНИЕ
Рис. 1. (А) Вид с боковой стороны на эмбрион мыши на стадии 4-6 сомитов. Очерченный район соответствует анализируемой области; cm - кардиальная мезодерма. (С) Передняя половина 8-сомитного эмбриона. Обозначения: 1 - кардиальная мезодерма; 2 - энтодермальная губа; 3 - средняя кишка; 4 - пунктирная стрелка показывает, что энтодерма передней кишки находтся позади клеток кардильной мезодермы.
Из всех исследований по генной инактивации у мыши до сих пор не обнаружено ни одной мутации, которая блокировала бы первоначальное развитие этих тканей печени и поджелудочной железы. Опыты с использованием культур эксплантатов привели нас к предположению, что рано на стадии сомитов неспособность продолжения развития (дефолт) по программе вентрального домена энтодермы передней кишки приводит к развитию поджелудочной железы. Культивированные кусочки энтодермы, выделенные с внешней поверхности передней кишки, не способны начать экспрессию генов поджелудочной железы, т.е. сами по себе условия культивирования обычно не индуцируют развитие поджелудочной железы. Из-за неспособности клеток вентральной энтодермы на стадии сомитов развиваться по программе поджелудочной железы происходит их переход на путь развития печени под влиянием находящейся вблизи кардиальной мезодермы (Рис. 2B). В этот период эмбриогенеза кардиальная мезодерма передвигается от латеральных доменов к средней линии, приводя к образованию кардиальной трубки. Мы предполагаем, что энтодерма, располагающаяся в стороне от образующейся кардиальной трубки и на губе, соединяющей переднюю кишку со средней (Рис.1C, 2B), находится достаточно далеко от кардиальной мезодермы, создавая условия для развития поджелудочной железы. Остается определить, существует ли определенный порог концентрации FGF, определяющий границу, которая отделяет область установления одного пути развития от области установления другого пути развития. Однако, дальнейшие морфогенетические перемещение и разрастание образующихся тканей приводит к четкому отделению печени от вентральной части поджелудочной железы (Рис.2C).
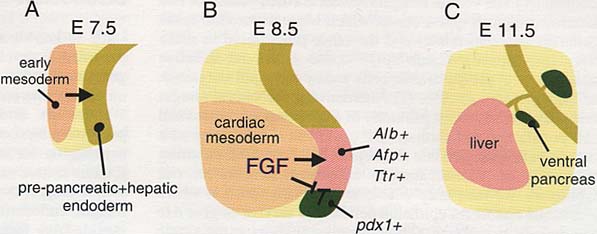
Рис. 2. Предполагаемые фазы раннего развития вентральной части поджелудочной железы. (А,В) Диаграммы сагитального сечения области вентральной части передней кишки. (А) Формирование раннего паттерна при воздействии мезодермы (стрелка) помогает создать бипотенциальность энтодермы передней кишки в отношении поджелудочной железы и печени. (В) Сигнал FGF от кардиальной мезодермы переводит клетки проксимальной энтодермы с развития по пути поджелудочной железы на путь развития печени (область окрашена розовым цветом), что доказано различиями в экспрессии обозначенных генов. Энтодерма, расположенная не в непосредственной близости к кардиальной мезодерме, например, на губе, которая соединяет переднюю кишку со средней, развивается в поджелудочную железу(область окрашена темнозеленым цветом). Тогда как кардиальная мезодерма обладает положительным эффектом на индукцию печени (стрелка), механистически она ,по-видимому, блокирует внутреннюю программу развития поджелудочной железы (barred перечеркнутые линии) и тем самым инициирует развитие печени в вентральной части энтодермы передней кишки. (С) Дальнейшие сигналы и морфогенез приводят к различию органов.
Рис. 2. Предполагаемые фазы раннего развития вентральной части поджелудочной железы. (А,В) Диаграммы сагитального сечения области вентральной части передней кишки. (А) Формирование раннего паттерна при воздействии мезодермы (стрелка) помогает создать бипотенциальность энтодермы передней кишки в отношении поджелудочной железы и печени. (В) Сигнал FGF от кардиальной мезодермы переводит клетки проксимальной энтодермы с развития по пути поджелудочной железы на путь развития печени (область окрашена розовым цветом), что доказано различиями в экспрессии обозначенных генов. Энтодерма, расположенная не в непосредственной близости к кардиальной мезодерме, например, на губе, которая соединяет переднюю кишку со средней, развивается в поджелудочную железу(область окрашена темнозеленым цветом). Тогда как кардиальная мезодерма обладает положительным эффектом на индукцию печени (стрелка), механистически она ,по-видимому, блокирует внутреннюю программу развития поджелудочной железы (barred перечеркнутые линии) и тем самым инициирует развитие печени в вентральной части энтодермы передней кишки. (С) Дальнейшие сигналы и морфогенез приводят к различию органов.
Существует много доказательств о происхождении и эндокринной, и экзокринной линий из единственной панкреатической стволовой клетки, а гепатоцитов и клеток желчного протока - из одного типа печеночных стволовых клеток или гепатобластов(Shiojiri, 1984; German et al., 1988). Мы ограничились рассмотрением первоначальной спецификации поджелудочной железы, показывая ,что выделенная энтодерма может давать начало клеткам, экспрессирующим маркеры ,характерные для эндокринной линии (Pdx1, ngn3, Isl1, BETA2/neuroD, glucagon и insulin), и инициальный р48 маркер экзокринной линии, не акцентируя внимание на дальнейшем развитии экзокринной линии. В наших исследованиях домен экспрессии Pdx1 близко окружал домен экспрессии HNF3бета, являющегося маркером для дефинитивной энтодермы, и клетки большинства выделенных кусочков энтодермы вентральной части передней кишки были компетентны к экспрессии панкреатического маркера . Несмотря на то, что даже небольшое количество клеток фибробластов, присутсвовавших в культурах, могло бы помочь установлению программы развития поджелудочной железы, некоторые культуры клеток энтодермы были компетентны к инициации развития печени, демонстрируя бипотенциальность. В прежних работах мы обнаружили, что участки этой энтодермы в присутствии клеток кардиальной мезодермы также начинают экспрессировать Ttf1, ранний транскрипционный фактор в развитии щитовидной железы и легких(Mizuno et al., 1991; Lazzaro et al., 1991; kimura et al.,1996), но связь между экспрессией Ttf1, Pdx1, доменами печени и сердца пока еще не определена.
Невозможность развития по пути поджелудочной железы на ранней стадии сомитов в вентральной энтодерме на Е8,5 (день появления вагинальной пробки рассматривался как Е0,5) вероятно отражает предшествующий патернинг на пресомитных стадиях (Рис.2A). Сразу после гаструляции, вентролатеральная энтодерма окружена висцеральной (splanchnic)мезодермой. Wells и Melton (2000) недавно показали, что при культивировании энтодерма на пресомитной стадии от Е7,5 развивается в направлении к Pdx1- положительной стадии только в присутствии фрагменов мезодермы и эктодермы. Ни FGF, ни BMP, никакой другой из тестированных факторов не способен индуцировать экспрессию Pdx1 в изолированной Е7,5 энтодерме. Вместе взятые данные показывают, что для невозможности развития по программе поджелудочной железы на стадии Е8,5 необходимы другие, еще не определенные сигналы от других зародышевых слоев на более ранних стадиях.
Способность кардиального сигнала FGF инициировать гепатогенез(Jung et al., 1999) можно интерпретировать как разрешение невозможности развития по программе печени или активирование генов печени в клетках, уже предопределенных к этому пути развития. Полученные данные отвергают эти возможности, так как мы обнаруружили, что невозможность развития для энтодермы вентральной части передней кишки относится к программе поджелудочной железы и, что в этом случае кардиальный сигнал FGF переводит клеточную популяцию на другой путь развития. Отсутствие действия кардиальной мезодермы или сигнала FGF на жизнеспособность и рост клеток энтодермы, расположенной проксимально, подтверждает концепцию, что сигнал FGF инициирует дифференциацию печени в клеточной популяции вентральной энтодермы. Эти данные объясняют некоторые курьезные наблюдения в наших прежних работах. Gualdi et al.(1996) и Jung et al., (1999) обнаружили, что еще задолго до гепатогенеза в энтодерме вентральной части передней кишки экспрессируются на очень низком уровне гены печени для альфа-фетопротеина(Afp) и транстиретина(Ttr). В культуре клеток в отсутствии кардиальной мезодермы или сигнала FGF экспрессия быстро гаснет. Результаты этой работы объясняют отсутствие AFP и TTR в изолированной энтодерме отчасти выбором клетками типа развития в направлении поджелудочной железы, которое исключает экспрессию генов , характерных для печени.
Заключение о невозможности развития поджелудочной железы в клетках энтодермы вентральной части передней кишки у мыши и отклонение пути развития посредством воздействия FGF2 согласуется с результатами исследований развития энтодермы у Xenopus. Эксплантаты вегетативного полюса от эмбрионов стадии ранней бластулы, свободные от мезодермы, культивированные в условиях изоляции, инициируют экспрессию гомолога Pdx1, X1Hbox8 (Gamer and Wright, 1995). Несмотря на то, что дополнительные маркеры поджелудочной железы не анализировали, FGF2 сильно подавлял экспрессию X1Hbox8 подобно действию FGF2 на энтодерму вентральной части передней кишки у мыши. В другой работе изолированные эксплантанты клеток вегетативного полюса автономно активировали экспрессию генов X1Hbox8 и инсулина(Henrry et al., 1996). Таким образом, невозможность развития поджелудочной железы в вентральной части энтодермы и его подавление при действии FGF может быть эволюционно законсервировано. Это контрастирует с ситуацией в дорзальной энтодерме, в которой не происходит обвала (defolt) пути развития поджелудочной железы в условиях изоляции (Kim et al., 1997) и где очень низкие уровни FGF2 стимулируют экспрессию Pdx1 (Hebrok et al. 1998). На поздних стадиях развития сигнал FGF необходим для развития экзокринной линии(Miralles et al., 1999).
Дальнейшее различие между дорзальным и вентральным доменами передней кишки это наша находка, что при невозможности дальнейшего развития в вентральной части энтодермы не происходит экспрессии Shh; в сравнении с этим Hebrok et al.(1998) и нами обнаружено, что в изолированной дорзальной энтодерме Shh экспрессируется. В то время, как нотохорда подавляет экспрессию Shh в дорзальной части , позволяя поджелудочной железе развиваться in vitro( Hebrok et al.,1998; Kim and Melton, 1998), клетки кардиальной мезодермы индуцируют локальную экспрессию Shh, которая может служить для локального поддержания развития поджелудочной железы. В самом деле, мы обнаружили, что для подавления экспрессии Pdx1 в клетках энтодермы передней кишки достаточно воздействия на них Shh. Мы предполагаем, что наблюдаемая экспрессия Shh в вентральной части энтодермs передней кишки эмбрионов мыши происходит вследствие широкого взаимодействия поверхности энтодермы с кардиальной мезодермой, приводящим к индукции Shh в большей части вентральной энтодермы. Ген Shh важен для морфогенеза трахей легких и esophagus(Chiang et al., 1996; Litingtung et al., 1998). Комбинация нуль-аллеля Shh в гомозиготе с нуль-аллелем Ihh (indian hedgehog)в гетерозиготе вызывает усиление раннего ветвления вентральной части поджелудочной железы (Ramalho-Santos et al., 2000; Неbrok et al., 2000), что совпадает с тем, что в раннем развитии поджелудочной железы происходит подавление сигналов Shh. Однако, эктопически в пределах домена экспрессии гена Pdx1 ген Shh подавлял, но не предотвращал развитие поджелудочной железы у эмбрионов мыши, по-видимому, из-за вмешивающегося процесса дифференциации расположенной рядом мезенхимы (Apelqvist et al., 1997). Аналогичным образом, при совместном культивировании клеток энтодермы и кардиальной мезодермы мы обнаружили в мезенхимальных клетках, окружающих энтодермальный эпителий, экспрессировавший Shh, экспрессию Ptc-2 (Ptch2), являющегося рецептором для Shh, и экспрессию генов ответа (Responsive)(Motoyama et al.,1998; Carpenter et al.,1998). У гомозиготных мутантов Ptc-1(Patch1) на ранних стадиях снижена экспрессия генов поджелудочной железы(Hebrok et al., 2000). Необходимы дальнейшие работы для оценки сложной взаимосвязи между Shh, отвечающей мезенхимой и развитием поджелудочной железы.
Несмотря на то, что в кардиальной мезодерме эмбриона мыши во время инициации развития печени и поджелудочной железы происходит индукция и FGF1, и FGF2 , инактивация любого из этих генов не мешает развитию кишки(Ortega et al., 1998; Dono et al., 1998; Miller et al., 2000). Kардиальная мезодерма также экспрессирует FGF4 (Zhu et al., 1996), FGF17(Maruoka et al., 1998), и на низком уровне FGF8 (crossley and Martin, 1998) и антагонист к общему сигналу FGF, ингибирующий гепатогенез in vitro (Jung et al., 1999); таким образом кардиальные сигналы являются высоко избыточными. Общее эмбриологическое происхождение вентральной части поджелудочной железы и печени может отражаться в определенных эволюционых и патологических состояниях. Примитивные беспозвоночные такие, как молюски имеют единственный орган гепатопанкреас, выполняющий функции обоих органов( Hoar, 1975). У морской миноги каудальный панкреас явно развивается путем трансдифференциации клеток печеночного протока(Elliot and Youson, 1993). Разделение печени и вентрального панкреаса в разные органы может эволюционировать(происходить) из общей клеточной популяции подобно тому, как описано здесь, а дорзальный панкреотический зачаток мог возникнуть в ходе поздней эволюционной адаптации.
У крыс, содержавщихся на определенной диете, которая вызывает повреждения в поджелудочной железе, в последней развивались кластеры гепатоцитов(Rao et al., 1989; Dabeva et al.,1995 Dabeva et al., 1997). Также трансгенная сверхэкспрессия FGF в поджелудочной железе взрослых мышей приводит к появлению гепатоцитов в островках (Krakowski et al., 1999). Более поздние находки определили роль сигнала FGF в инициации дифференциации печени, но не поджелудочной железы. В этих случаях неизвестно, появляются ли панкреатические клетки путем трандифференциации клеток пакреатического протока или путем альтернативной спецификации популяции еще неопределившихся стволовых клеток, которая могут быть ассоциирована с компартментом протока(Slack, 1985; Gu and Sarvetnick, 1993; Krizik et al., 1999). Тем не менее, появление клеток печени в поджелудочной железе взрослых особей иллюстрирует соотношение спецификации этих тканей в развитии, обнаруженное нами, и дает новый точку зрения относительно того, как может контролироваться дифференциация клеток, что может быть использовано для клеточной терапии заболеваний.