Щелевые соединения выполняют множество разных ролей в различных тканях. 3 разных профиля проницаемости обнаружено для щелевых соединений в сетчатке, что предполагает участие разных типов коннексинов. Наиболее проницаемыми для больших молекул были щелевые соединения от А-типа горизонтальных клеток. Проницаемость щелевых содеинений была одинакова в двух типах амакринных клеток и В-типе горизонтальных клеток. Самая низкая проницаемость была найдена в щелевых соедниениях между конусными биполярными клетками и AII амакринными клетками, с которыми они купированы.
Получены доказательства того, что между членами всех пяти основных классов нейронов сетчатки существуют щелевые соединения. В этот процесс вовлечены гетеротипические коннексоны и множественные типы коннексинов.
| Rod Pathways in the Mammalian retina Use Connexin 36 MILLS S.L, O'BRIEN J.J, LI W, O'bRIEN О, MASSEY S.C THE JOURNAL OF COMPARATIVE NEUROLOGY V. 436. N 3. P. 336-350 (2001) |
Получены доказательства того, что между членами всех пяти основных классов нейронов сетчатки существуют щелевые соединения. В этот процесс вовлечены гетеротипические коннексоны и множественные типы коннексинов.
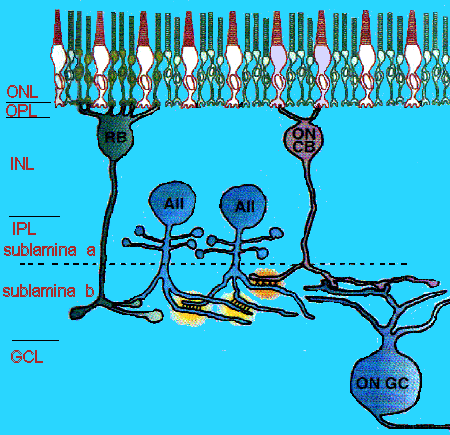
Рис. 1. Схематическая диаграмма роли амакринных клеток AII в зрительном пути от фоторецепторов палочек. Каждая AII амакринная клетка формирует щелевые соединения не только с соседними AII клетками, но и с ON конусными биполярными клетками. Эти sign-preserving щелевые соединения с биполярными клетками распространяют сигналы от палочковых биполярных клеток, которые деполяризуют свет, в ON конусные биполярные клетки, которые такжже деполяризут свет. Sign-inverting ингиибрующие синапсы от AII амакринных клеток к OFF конусным биполярным клеткам наблюдаются в субламине (не показаны), они обеспечивают распределение сигнала с соотвествующей гиперполяризацией света в OFF канал. Необходимые для разделения палочковых и колбочковых ганглиолярных клеток сигналы отсуствуют. ONL, наружный ядерный слой; OPL, наружный плексиформный слой; INL, внутренний ядерный слой; IPL, внутренний плексиформный слой; GCL, слой ганглиолярных клеток; RB, палочковые биполярные клетки; CB, конусные биполярные клетки; GC, ганглиолярная клетка.
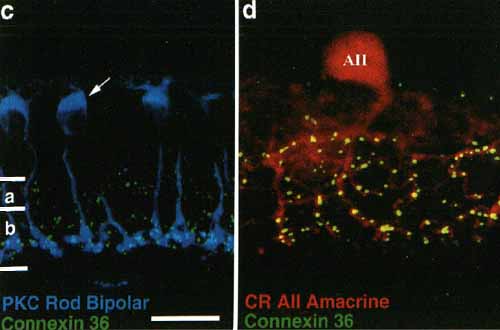
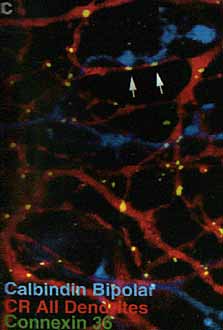
Рис.3. с. Окраска палочковидных биполярных клеток (голубые) с помощью антител к протеин киназе С (РКС) демонстрирует колокализацию с диффузным Сх36 иммуноокрашиванием (зеленое). Полоска = 20 µm. Строгая ко-локализация Сх36 иммунореактивности (зеленое), обнаруживаемой на отростках в подслое AII амикринных клеток, окрашенных анти-калретинином (CR, красное). Светлая Сх36 иммунореактивность в подслое а не ассоциирована с AII лобулярными придатками. Полоска = 10 µm.
Рис. 8.с. Окрашенные калретином отростки AII амакринных клеток (красные)постоянно обнаруживаются около отростков биполярных клеток (крашены антителами к калбиндину, голубые), между ними видны Сх36 иммунореактивные точки (светло-зеленые).
AII амакринные клетки образуют гомологичные щелевые соединения с соседними AII амакринными клетками и гетерологические щелевые соединения с ON конусными биполярными клетками. AII амакринные клетки являются критическими интернейронами в зрительном пути от палочек (rod pathways vision) у млекопитающих. Сигналы от колбочек (cones) непосредственно подразделяются на "ON" (ВКЛ) и "OFF" (ВЫКЛ) каналы конусных биполярных клеток, которые реагируют на свет с противоположной полярностью (Рис. 1). Однако имеется только один тип палочковых биполярных клеток, которые обеспечивают "ON" ответ. Палочковые биполярные клетки не образуют синапсов непосредственно с ганглиолярными клетками. Вместо этого сигналы от палочек попадают как в ON, так и OFF каналы посредством AII амакринных клеток. AII обеспечивают это за счет щелевых соединений с ON конусными биполярными клетками и за счет sign-inverting ингибирующих синапсов на OFF конусных биполярных клетках. Купирование между AII амакринными клетками, по-видимому, улучшает соотношение сигнал/шум на уровне слабого света, тогда как модулирование связи между AII/конусными биполярными клетками имеет целью передачу зрительного сигнала от палочек к колбочкам.
Было установлено, что в сетчатке кроликов антитела к коннексину 36 сильно метят отростки AII амакринных клеток. Установлено, что Сх36 первоначально локализуется дендритных пересечениях между перекрывающимися AII амакринными клетками. Следовательно Сх36 участвует в образовании гомотипических щелевых соединений между парами AII амакринных клеток. Сх36 обнаружен также в местах контактов между AII и, по-видимому, ON конусными биполярными клетками, в местах образования гетеротипических щелевых соединений. Известно, что щелевые соединения между AII и ON конусными биполярными клетками значительно менее проницаемы для крупных трассеров, чем каналы между соседними AII клетками. Характерным для каналов Сх36 являются их слабые voltage-gating свойства. Следовательно, resting мембранный потенциал AII амакринных клеток скорее более 20 mV и более негативный, чем у ON конусных биполярных клеток. Метка Сх36 в других областях сетчатки не связана с AII амакринными, а некими другими клетками.
Отсутствие ассоциации Сх36 с горизонтальными клетками и S1.S2 амакринными клетками указывает на присутствие еще неидентифицированных типов коннексинов в этих клетках. В сетчатке рыбок данио, напр., найдено 4 типа коннексинов, три из них ранее не были известны. Предполагается существование не менее 6 типов коннексинов в сетчатке кроликов.
Cx36 выявлен в AII клетках также в сетчатке мышей и крыс ( Feigenspan et al., J.Neurosci. V.21. P. 230. 2001) и в местах контактов их с ON конусными биполярными клетками.
|
Coupling From AII Amacrine Cells to ON
Cone Bipolar Cells Is Bidirectional E. BRADY TREXLER, WEI LI, STEPHEN L. MILLS, AND STEPHEN C. MASSEY THE JOURNAL OF COMPARATIVE NEUROLOGY 437:408-422 (2001) |
Первые результаты показывали, что AII/bipolar щелевые соединения позволяют проходит трассеру только в одном направлении. Это контрастировало с результатами физиологических иследований, которые указывали на то, что свет, адаптированный в сигналы м. проходить от ON конусных биполярных клеток в сеть AII амакринных клеток. В данной работе было показано, что различные ON и OFF биполярные клеткииногда аномально купируются с сетью горизонтальных клеток А-типа.
Эти довольно редкие сбытия представляют собой минорные ошибки развития.
Однако, это делает возможным заплнить Neurobiotin конусные биполярные клетки, не вводя в них микроэлектроды. В этих условиях Neurobiotin распространяется от ONконусных биполярных клеток в соседние AII амакринные клетки. Эти результаты указывают на то, что щелевые соединения между AII/bipolar клетками позволяют трассеру перемещаться в обоих направлениях.
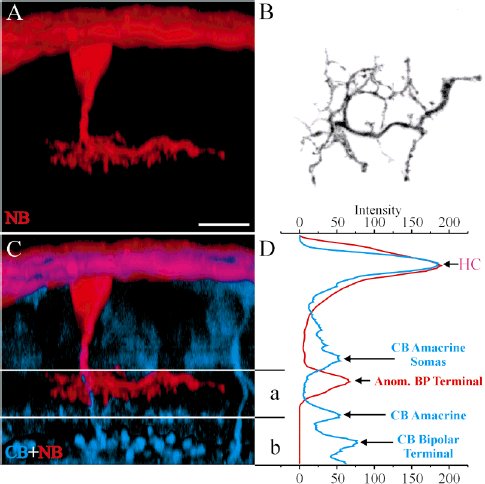
Fig. 5. An example of anomalously coupled OFF bipolar cell de- fined by the calbindin amacrine cell as a marker of the center of the IPL. A: A Z-axis projection of Neurobiotin labeling in optical sections ranging from the OPL to the bottom of the IPL depicts an anomalously coupled bipolar cell. B: An X-Y projection of the same cell shows the detailed structure of the axon terminal. C: A double-labeled confocal image with Neurobiotin coded in red and calbindin in blue. Calbindin labels horizontal cells, one type of ON bipolar cell with axon terminal at the lower part of IPL, and one type of wide field amacrine cell whose dendrites ramify in the middle of IPL. This bipolar cell’s axon terminal is located approximately 5 mm above the band of calbindin amacrine cell. Thus, it is an OFF bipolar cell. D: The average color intensity for each horizontal line of the image in C was analyzed. The plot was scaled to coincide with the image in C. The blue curve represents the calbindin signal. In the IPL there are two peaks, one in the middle of IPL, which is from the calbindin amacrine cells, and the other at the bottom that is from the terminals of calbindin bipolar cells. The red curve representing the Neurobiotin signal has two peaks. The large one at the top corresponds to the horizontal cells and the lower one in the IPL represents the axon terminal of the coupled bipolar cell, which is slightly above and overlapping the blue peak of calbindin amacrine cell. NB, Neurobiotin; CB, calbindin; Anom., anomalous; BP, bipolar; IPL, inner plexiform layer. Scale bar 5 10 mm.
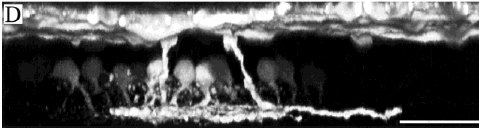
Рис.7 D: A Z-axis projection of the same optical sections shows Neurobiotin labeling from the OPL to the bottom of the IPL. The A-HC network is at the top. The two anomalously coupled bipolar cells ramify at slightly different levels near the bottom of sublamina b. The other Neurobiotin-labeled cells have morphology characteristic of AII amacrine cells. There are no direct connections between these AII amacrine cells and the Neurobiotin-filled A-type horizontal cell network. Presumably, the AIIs were filled with Neurobiotin by means of the anomalously coupled bipolar cells. INL, inner nuclear layer; OPL, outer plexiform layer. Scale bars 5 30 mm
Несколько линий доказательств указывают на то, что щелевые соединения между AII и конусными клетками м. отличаться от AII/AII щелевых соединений. Так, при ЭМ II/bipolar щелевяе соединения асмимметричны. В дополнение к тому, что большме биотинилированные трассеры проходят довольно плохо через AII/bipolar щелевые соединения, т.е. они меньше, чем AII/AII щелевые соединения. Купирование в AII/AII сети тонко контролируется с помощью dopamine, тогда как купирование AII/bipolar клеток модулируется с помощью NO/cGMP-зависимого пути. Это согласуется с физиологическими данными, что большие фотооптические ON реакции, записываемые с AII амакринных клеток, скорее всего поступающие с ON конусных биполярных клеток через AII/bipolar щелевые соединения, м.б. модулированы с помощью NO/cGMP. Напротив, адаптированный к темноте ответ AII, который отражает синаптический импульс от палочковых биполярных клеток, не испытывают влияния со стороны NO/cGMP. Наконец, AII/bipolar купирование по иному редуцируется carbenoxolone.
Fig. 5. An example of anomalously coupled OFF bipolar cell de- fined by the calbindin amacrine cell as a marker of the center of the IPL. A: A Z-axis projection of Neurobiotin labeling in optical sections ranging from the OPL to the bottom of the IPL depicts an anomalously coupled bipolar cell. B: An X-Y projection of the same cell shows the detailed structure of the axon terminal. C: A double-labeled confocal image with Neurobiotin coded in red and calbindin in blue. Calbindin labels horizontal cells, one type of ON bipolar cell with axon terminal at the lower part of IPL, and one type of wide field amacrine cell whose dendrites ramify in the middle of IPL. This bipolar cell’s axon terminal is located approximately 5 mm above the band of calbindin amacrine cell. Thus, it is an OFF bipolar cell. D: The average color intensity for each horizontal line of the image in C was analyzed. The plot was scaled to coincide with the image in C. The blue curve represents the calbindin signal. In the IPL there are two peaks, one in the middle of IPL, which is from the calbindin amacrine cells, and the other at the bottom that is from the terminals of calbindin bipolar cells. The red curve representing the Neurobiotin signal has two peaks. The large one at the top corresponds to the horizontal cells and the lower one in the IPL represents the axon terminal of the coupled bipolar cell, which is slightly above and overlapping the blue peak of calbindin amacrine cell. NB, Neurobiotin; CB, calbindin; Anom., anomalous; BP, bipolar; IPL, inner plexiform layer. Scale bar 5 10 mm.
Рис.7 D: A Z-axis projection of the same optical sections shows Neurobiotin labeling from the OPL to the bottom of the IPL. The A-HC network is at the top. The two anomalously coupled bipolar cells ramify at slightly different levels near the bottom of sublamina b. The other Neurobiotin-labeled cells have morphology characteristic of AII amacrine cells. There are no direct connections between these AII amacrine cells and the Neurobiotin-filled A-type horizontal cell network. Presumably, the AIIs were filled with Neurobiotin by means of the anomalously coupled bipolar cells. INL, inner nuclear layer; OPL, outer plexiform layer. Scale bars 5 30 mm
Несколько линий доказательств указывают на то, что щелевые соединения между AII и конусными клетками м. отличаться от AII/AII щелевых соединений. Так, при ЭМ II/bipolar щелевяе соединения асмимметричны. В дополнение к тому, что большме биотинилированные трассеры проходят довольно плохо через AII/bipolar щелевые соединения, т.е. они меньше, чем AII/AII щелевые соединения. Купирование в AII/AII сети тонко контролируется с помощью dopamine, тогда как купирование AII/bipolar клеток модулируется с помощью NO/cGMP-зависимого пути. Это согласуется с физиологическими данными, что большие фотооптические ON реакции, записываемые с AII амакринных клеток, скорее всего поступающие с ON конусных биполярных клеток через AII/bipolar щелевые соединения, м.б. модулированы с помощью NO/cGMP. Напротив, адаптированный к темноте ответ AII, который отражает синаптический импульс от палочковых биполярных клеток, не испытывают влияния со стороны NO/cGMP. Наконец, AII/bipolar купирование по иному редуцируется carbenoxolone.
Если AII/AII щелевые соединения гомотипичны, то AII/bipolar щелевые соединения м.б. гетеротипичными, т.е. разные коннексины формируют полуканальцы на каждой стороне щелевого соединения.
AII амакринные клетки экспрессируют коннексин 36, но идентичность коннексинов ON биполярных клеток еще не установлена. Возможно, что они также экспрессируют Сх36, но он по-другому модулируется со стороны биполярных клеток, обусловливая их ассмметрию. Другие коннексины также экспрессируются в сетчатке. Если эти щелевые соединения действительно гетеротипические, то возможно, что коннексины биполярных клеткок очень чувствительны к transjunctional voltage, введение электродов вызывает деполяризацию.