Так как эктопическая Vg1 обусловливает инверсию оси LR только тогда, когда вводится в правые (но не левые) латеральные вегетативные клетки, то очевидно, что активация Vg1 сигнального пути обычно пространственно ограничена левой латеральной областью бластулы Рис.1.
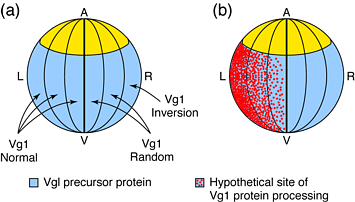
Fig. Inversion of the left-right (LR) axis by Vg1. (a) Vg1 precursor protein is expressed by left and right vegetal cells of the early Xenopus blastula (blue). Ectopic expression of mature Vg1 protein in right, but not left, vegetal cells results in altered LR situs. Microinjection of Vg1 RNA or cDNA (arrows) in right dorsovegetal cells of the 16-cell blastula results in heterotaxia; expression by the right ventral-lateral lineage results in subsequent inversion of the LR axis. (b) Because ectopic expression of mature Vg1 affects LR development only when introduced to the right side of the embryo, it is hypothesized that the activity of Vg1 ligand normally is restricted to cells of the left vegetal region, perhaps through a mechanism involving asymmetrical generation of mature Vg1 from its precursor protein (red)
В соответствии с этой гипотезой доминантно-негативный TGF-β рецептор , ингибирующий Vg1 signaling , вызывает инверсию сердца и органов, когда экспрессируется в левых (но не правых) латеральных вегетативных клетках 13 14. По-видимому, ось LR формируется или во время или после стадии, на которой возможны экспериментальные нарушения. У эмбрионов кур ось LR также ,по-видимому, закладывается до гаструляции. На стадии 4 ткани латеральнее узла уже определили свое положение относительно LR благодаря индуктивным взаимодействиям 15. Однажды индуцированный в отношении LR identity, узел затем управляет развитием LR путем возвращения сигналов назад в соседние ткани. Если узел оказывается ревертированным в отношении своей LR спецификации (e.g. through surgical rotation of the stage 6 node) , то LR позиционная информация узла является доминантной по отношению к позиционной информации, перед этим имевшейся в тканях латеральнее узла.
Ткани, расположенные по бокам от организатора/узла, и способные влиять на последующую асимметрию , включая LR спецификацию организатора/узла названы лево-правосторонним координатором `left-right coordinator' (LRC) у Xenopus14. В этой модели активность LRC в левых латеральных вегетативных клетках, возможно опосредуемая передачей сигналов Vg1 , обусловливает `left identity' в этих клетках. Клетки же на противоположной стороне эмбриона вынуждены адоптировать `right identity'. Эктопическая экспрессия зрелого Vg1 правыми латеральными клетками преодолевает эндогенную передачу сигналов Vg1 на левую сторону, в результате размещения эктопического LRC справа.
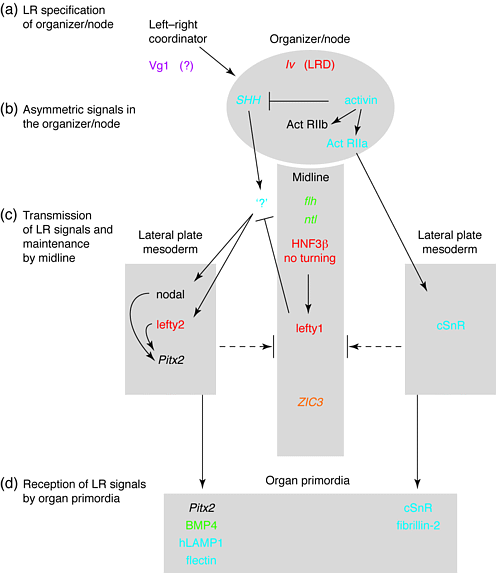
Fig. Summary of inductive interactions in the left-right (LR) developmental pathway. (a) LR coordinator activity generates LR identity in tissues lateral to the organizer/node. By an undefined mechanism, these tissues in turn elicit a cascade of asymmetrical gene expression in the organizer/node. (b) Once specified for LR identity, the node signals back to its surrounding tissues to elicit asymmetrical gene expression in paraxial and lateral plate mesoderm. (c) Following regression of the organizer/node, the notochord and floor plate function to maintain asymmetry of gene expression to either side of the midline and to prevent diffusion of molecules from one side of the embryo to the other (dashed arrow with bar). (d) Transmission of LR patterning cues from the lateral plate mesoderm results in asymmetrical gene expression by organ primordia. Genes and proteins that are expressed at stages coincident with these inductive processes are represented by color to indicate the organism in which they were experimentally determined to function in the LR pathway: frog (purple); chick (blue); mouse (red); zebrafish (green); human (orange); or more than one organism (black). Arrows and bars represent positive and negative regulatory interactions between genes, but do not necessarily imply direct interactions. Unidentified signals are indicated by `?'.
Таким образом, эксп. нарушения , ведущие к инверсии или рандомизации LR оси могут обусловливаться взаимодействием с LRC активностью и/или с LR спецификацией узла.
Asymmetric signals in the organizer/node
У эмбрионов кур обмен позиционной информацией между узлом и соседними тканями обеспечивается путями передачи сигналов activin и shh 15 16. Этот сигнальный каскад - первая идентифицированная молекулярная асимметрия у ранних эмбрионов (see Refs 134 for review), причем право-сторонняяя экспрессия активина β B в узле индуцирует асимметричную экспрессию рецептора активна (ActRIIa) и shh (Fig 2b). Эктопическая активация или ингибиция этого пути обусловливает рандомизацию петлеобразования сердца и неправильную экспрессию нижестоящих мишеней этого сигнального каскада (e.g. nodal, see below) (Refs 151617). Неизвестный вторичный сигнал(ы) необходимы пути activin/shh , так как эктопический белок SHH индуцирует экспрессию nodal в эксплантированной мезодерме латеральной пластинки только если параксиальные ткани включены в культуру 15. Однако у других позвоночных доказательства действия эндогенных activin и shh путей не находят подтверждения, возможно что иные, но сходные с activin и shh оперируют передачей сигналов закладывающих ось LR у джругих видов . Возможно также, что ранние ступени закладки асимметрии не законсервированы.
Transmission of LR patterning by lateral plate mesoderm and maintenance of asymmetry by the midline
Регулируемая асимметричная экспрессия генов в мезодерме латеральной пластинки (LPM) является первым четким указанием на законсервированный механизм формирования LR паттерна у позвоночных (Fig 2c). Сходная с TGF-β молекула nodal экспрессируется в левой LPM у всех позвоночных . Сложный паттерн экспрессии nodal, варьирующий от правосторонней локализации, билатеральной локализации или до отсутствия экспрессии выявляется у iv/iv мышей 26 and правосторонняя экспрессия у inv/inv мышей 2326. Аномальная правосторпонняя экспрессия nodal in Xenopus25 и эмбрионов кур 17 обусловливает рандомизацию петлеобразования . Presumably, bilateral (or absent) nodal expression represents an ambiguous cue for LR orientation, hence resulting in stochastic alignments along the LR axis.