ЦИТОФЕНОТИП НЕЙРОНА
Формирование нервноклеточного цитофенотипа осуществляется в рамках трех категорий корреляций: геномных ( внутренняя генетическая программа спецификации), морфогенетических (связи между клеточными ассоциациями в периодаы гисто- и органогенеза) и эргонтических (терминальная клеточная дифференцировка как эффект функционирования) (Корочкин, 2000).
ЦИТОФЕНЕТИКА-1
"Внутренняя программа" развития нейрона, обнаруживаемая в становлении определенного конкретного цитофенотипа, регистрируется как специфическая последовательность появления цитогенотипических и цитофенотипических различий между системами и группами клеток. Так, уже у 11-12 недельных зародышей человека проявляется тенденция периферических парасимпатических нервных клеток пищеварительной трубки к развитию по пути интенсификации окислительно-восстановительных процессов (высокая активность сукцинатдегидрогеназы и цитохромоксидазы). Напротив, в периферических симпатических нейронах отмечается накопление гликогена и "отсроченная" по времени активация сукциноксидазной системы. Кроме этого в клетках парасимпатических сплетений пищеварительной трубки постепенно нарастает активность кислой фосфатазы и эстеразы, в то время как для дифференцирующихся симпати ческих нервных клеток пограничного ствола более характерно нарастание активности щелочной фосфатазы. Обнаруживаются и другие белковые различия. Очевидно, это является следствием активации новых групп генов, ответственных за синтез определенных белковых соединений. В дальнейшем однородная популяция парасимпатических (и симпатических) нейронов распадается на клеточные догелевские типы по дивергентному принципу. Предположительно чувствительные клетки II типа Догеля отличаются при этом высокой активностью моноаминооксидазы. В то же время среди клеток I (и возможно, II) типа Догеля существует немалое разнообразие по их эргичности. Наряду с холинергическими среди них встречаются пурин-, холецистокинин-, соматостатинергические и др. Возникновение многочисленных развновидностей клеток I и II типов Догеля вероятно обусловлено различным пространственным расположением этих клеток, например, в ганглии. Однако все особенности, вследствие которых вегетативные нервные клетки иногда даже отклоняются от классических догелевских типов, касаютя главным образом морфологической характеристики клеток и не затрагивают их гистохимических параметров. Поэтому автор (Корочкин, 2000) относит эти особенности к цитофенотипическим. нервные клетки, связанные цитогенотипической общностью, как правило, обладают и близмкими реактивными свойствами. В частности, реактивные изменения I типа Догеля характеризуются большей частью увеличением поверхности дендритических ламелл, Кюгель-феноменом, образованием пластинчатых выростов. Для клеток II типа Догеля более характерно развитие корзинчатых сплетений и так называемых "окон", причем они по своим реактивным свойствам( а иногда и морфологическим признакам) порою обнаруживают некоторое сходство с чувствительными нейронами спинальных ганглиев. Это может свидетельствовать о наличии у тех и других сходныъх функционально активных генов.
Как созревание, так и функционирование нервных клеток автономной нервной системы осуществляется группами, ансамблями ( то, что теперь в ЦНС называют "модулями"). Интересно, что и в ходе патологического процесса нервные клетки болеют и гибнут гнездами. В целом складывается совершенно определенное впечатление о необходимости взаимодействия какого-то количества клеток для нормального течения процесспа дифференцировки ( феномен минимальной массы).
Согласно схеме Лаврентьева, имеется определенная закономерность в количественном соотношении между клетками I и II типа Догеля в разных участках пищеварительной трубки млекопитающих. Это объясняется индуцирующим воздействием преганглиолярных волокон на нейробласты интрамуральных сплетений пищеварительного тракта. Верхние отделы этого тракта в избытке обеспечиваются волокнами блуждающего нерва, количество которых уменьшается по мере продвижения в каудальном направлении. (градиент). Нижние отделы ( нижние сегменты толстой кишки, прямая кишка) "насыщаются" преганглиолярными парасимпатическими волокнами, которые образованы n.pelvicus и количество которых уменьшается в краниальном направлении (каудокраниальный градиент).
Можно предполагать, что нейробласты, вступившие в контакт с преганглиолярными волокнами, дифференцируются в в I догелевский тип, а нейробласты, оставшиеся "свободными", развиваются во II тип Догеля. Однако отростки клеток II типа растут не в случайных направлениях, а часто контактируют с дендритами соседних нейронов I типа. Создается впечатление, что около каждой дифференцирующейся нервной клетки I типа складывается своеобразное морфогенетическое поле, в направление которого растут отростки детерминированных во II тип Догеля нейробластов. В подобных взаимодействиях из физических факторов могут иметь значение электрические или митогенетические ( ультрафиолетовое излучение активно растущих тканей). Что касается химических факторов, то нельзя исключить возможности выделения определенных активных веществ преганглиолярными волокнами и их околоклеточными терминальными разветвлениями (ферментативная активность которых в период роста повышена). Возможно, это - медиаторы, ибо они, как известно, при воздействии на нейробласты, ещн лишенные аксона и дендритов, стимулируют образование многочисленных клеточных отростков.
ЦИТОФЕНЕТИКА-2
Терминальная дифференцировка нервной клетки, "оформление" ее цитофенотипа происходит в тесной связи с началом ее функционирования, с включением ее в тот или иной нейронный ансамбль в качестве составного компонента. В частности, специфическая архитектура нисслевской субстанции в перикарионе складывается окончательно лишь после того, как удается зарегистрировать установление функциональных связей соответствующей нервной клетки. Например, на телах нейронов интрамурального аппарата развиваются синапсы конечного типа (на 3-3.5 мес зародышевого развития в верхних отделах пищеварительной трубки плодов человека, на 3.5-4 мес в нижних ее отделах). С усложнением организации описанных синапсов ( у зародышей 3.5-4.5 мес в верхних отделах и у зародышей 4-4.5-5-мес - в нижних отделах пищеварительной трубки) нисслевское вещество начинает заполнять все тело крупных центральных вегетативных нейронов, в ядре обычно остается лишь одно ядрышко.
Если обратиться к симпатической нервной системе, то можно увидеть в основном ту же картину, хотя дифференцировка начинается несколько позднее. Как и в парасимпатическоей нервной системе, ей предшествует развитие преганглиолярных нейронов, в данном случае клеток боковых рогов спинного мозга. У зародышей 11-12 недель развития эти клетки обнаруживают первые признаки дифференцировки - рост ядра, появление отростков, усложнение нейрофибриллярного аппарата и возрастание его сродства к серебру, утрата равномерного распределения ДНК хромосом по ядру, перинуклеарное развитие нисслевких телец, накопление в ядрышке основных белков и т.д. Нервоноклеточные элементы ганглиев пограничного ствола грудной области вступают в фазу дифференцировки на 4-5 мес развития. Этой фазе предшествует интенсивное развитие интерцеллюлярного сплетения нервных волокон (у зародышей 3-3.5 мес).
Известно, что появление выраженной реакции сокращающегося кишечника человеческого зародыша на пилокарпин и адреналин совпадают по времени (9-12 неделя) с началом процессов дифференцировки нервноклеточных элементов ауэрбаховского сплетения и образования связей их отростков с окружающими тканями. Для нормального развития нервной клетки необходимы раздражения с периферии. Этот принцип применим также к вегетативным нейронам, и ускоренное увеличение размеров ядер дифференцирующихся нейронов вследствие усиленной функциональной нагрузки выявляется в сплетении пищеварительной трубки 6-мес плода.
Схема организации парасимпатического регуляторного нервного аппарата пищеварительной трубки.
Автор (Корочкин, 2000) предполагает наличие связи терминальных отростков эффекторных интрамуральных нейронов преимущественно с мышечными клетками пищеварительной трубки (рис).

Автор (Корочкин, 2000) предполагает наличие связи терминальных отростков эффекторных интрамуральных нейронов преимущественно с мышечными клетками пищеварительной трубки (рис).
Схема организации парасимпатического нервного аппарата пищеварительной трубки: 1-4 - мышцы ( 1 - ворсинок, 2 - t.muscularis mucosae, 3 - циркулярного слоя t. muscularis externa, 4 - продольного слоя t.muscularis externa), 5 - pl.submucosus, 6 - pl. infernus, 7 - ауэрбаховское сплетение, 8 - дорсальное ядро блуждающего нерва, 9 - спинальный ганглий. Нейроны I типа Догеля обозначены черным, а нейроны II типа Догеля - светлым. Стрелки указывают направление распространения нервного импульса.
Осуществление нейральной регуляции иных функций, например, выделение секрета, сооотносится с "выжиманием" последнего из клеток секреторного эпителия ( а также со слющиванием части этих клеток) обусловленных мышечными сокращениями ( в особенности сокращениями мышц слизистой оболочки).
ЦИТОФЕНЕТИКА-3
Под влиянием того или иного гормона активируется функция генетического аппарата ( усливается транскрипция) и тем самым ускоряется развитие или интенсифицируются функции соответствующей тканевой системы.
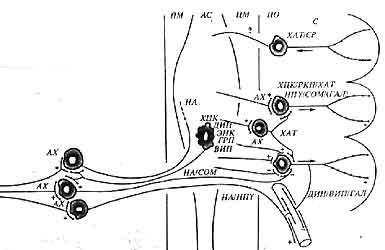
Схема нейронных типов мейсснеровского сплетения, их проекции, синаптических входов и физиологических эффектов (по Furness et al., 1988)
Схема нейронных типов мейсснеровского сплетения, их проекции, синаптических входов и физиологических эффектов (по Furness et al., 1988)
ПМ - продольные мышцы, АС - ауэрбаховское сплетение, ЦМ - циркулярные мышцы, ПО - подслизистая оболочка (submucosa), С - слизистая оболочка (mucosa), ХАТ - холинацетилтрансфераза, СР - субстанция Р, ХЦК - холецистокинин, ЗКП - родственный кальцитонину пептид, НПY - нейропептид Y, СОМ - соматостатин, ГАЛ - галанин, НА - норадреналин, ДИН - динорфин, ЭНК - энкефалин, ГРП - гастриновый релизинг-пептид, ВИП - вазоактивный интестинальный пептид, АХ - ацетилхолин
Клетки-предшественники симпатических нейронов способны выбирать один из двух путей развития - стать симпатобластами или хромафинными клетками. Предполагается, что это две фазы одного и того же пути и "задержака" на стадии хромафинной клетки подобна явлению неотении в мире живтоных. Оказалось также, что дифференцировка в направлении симпатобластов стимулируется геном фактора роста нервов (NGF), а дифференцировка в направлении хромаффинных клеток - глюкокортикоидами.
Очевидно, что генетические системы, контролирующие нейроэндокринные структуры, их развитие и функции, играют важную роль в детерминации процента нейронов с разной ергичностью и существенно влияют тем самым на специфический паттерн формирующихся нейронных ансамблей и особенности поведения, определяемые конструкцией этих ансамблей.