ЭНДОКАРДИАЛЬНЫЕ КЛЕТКИ И ТРУБКИ
Во время эпителизации кардиомиоцитов происходит выделение и популяции эндокардиальных клеток. Эндокардиальные клетки возникают из клеток вентральной спланхнической мезодермы (Рис.1)
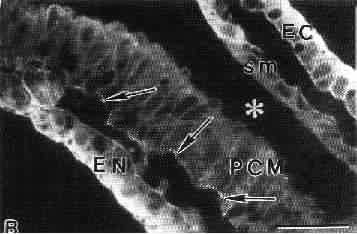
Распределение интегрина на базальной поверхности дифференцирующихся кардиомиоцитов после образования перикардиального целома у эмбрионов кур (ст.8). Это соответствует области высокой концентрации фибронектина. Эндотелиальные клетки, которые начинают отсортировываться от кардиомиоцитов на поверхности, обращенной к энтодерме также экспрессируют интегрин (Linask, 1992).
Обозначения: EC - эктодерма, sm - соматическая мезодерма, PCM - прекардиальные клетки, EN - энтодерма. Звездочкой обозначен перикардиальный целом (амионкардиальный пузырек). Стрелаками отмечены отдельные места экспресси интегрина.
Обозначения: EC - эктодерма, sm - соматическая мезодерма, PCM - прекардиальные клетки, EN - энтодерма. Звездочкой обозначен перикардиальный целом (амионкардиальный пузырек). Стрелаками отмечены отдельные места экспресси интегрина.
Они остаются мезенхимными, не эпителизируются и не становятся частью удлиняющегося компартмента кардиомиоцитов. Популяция клеток тесно связанных миокардиоцитов формируют дорсальный барьер для движения клеток, поэтому мезенхимные клетки могут отсортировываться только в вентральном направлении, к энтодерме (Рис.2)
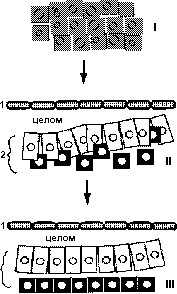
Рис.2.
Модель сортировки клеток кардиогенных областей (Linask et al., 1997).(I) На стадии 4-5 все клетки экспрессируют N-кадхерин и бета-катенин. Затем их экспрессия ограничивается фокусами , расположенными примерно посредине и отделяющими вентральную мезодерму (2) от дорсальной (1). Затем в этих фокусах возникают полости, которые сливаются и образуют перикардиальный целом, отделяющий соматическую мезодерму от спланхнической (II). Клетки вентральной мезодермы соединяются вместе с помощью N-кадхериновых адхерентных соединений (белые клетки). В процессе эпителизации популяция клеток, которая располагается более вентрально (черные клетки) начинают исключаться (II). После образования компартмента кардиомиоцитов презумптивные эндокардиальные клетки отсортировываются в направлении энтодермы (III).
I - латеральная мезодрема, включающая соматические и кардиальные предшественники, II - эпителизация миокардиоцитов (белые клетки) приводит в исключению эндокардиальных предшественников (черные клетки), III- эндокардипальные предшественники отсортировываются от вентраьных миокардиоцитов и дифференцируются в эндокардиальные тяжи
Модель сортировки клеток кардиогенных областей (Linask et al., 1997).(I) На стадии 4-5 все клетки экспрессируют N-кадхерин и бета-катенин. Затем их экспрессия ограничивается фокусами , расположенными примерно посредине и отделяющими вентральную мезодерму (2) от дорсальной (1). Затем в этих фокусах возникают полости, которые сливаются и образуют перикардиальный целом, отделяющий соматическую мезодерму от спланхнической (II). Клетки вентральной мезодермы соединяются вместе с помощью N-кадхериновых адхерентных соединений (белые клетки). В процессе эпителизации популяция клеток, которая располагается более вентрально (черные клетки) начинают исключаться (II). После образования компартмента кардиомиоцитов презумптивные эндокардиальные клетки отсортировываются в направлении энтодермы (III).
I - латеральная мезодрема, включающая соматические и кардиальные предшественники, II - эпителизация миокардиоцитов (белые клетки) приводит в исключению эндокардиальных предшественников (черные клетки), III- эндокардипальные предшественники отсортировываются от вентраьных миокардиоцитов и дифференцируются в эндокардиальные тяжи
Базируясь на экспрессии эндотелиального маркера перепела QH-1, определили начало появления клеток эндотелиальных предшественников на стадии 7+ (two somites) в задних частях билатеральных сердце-формирующих областей . Клетки, экспрессирующие антиген QH-1 были мезенхимными и располагались между мезодермальным эпителием кардиальных областей и энтодермой. С помощью конфокальной микроскопии выявлено асимметричное распределение QH-1 позитивных клеток между двумя кардиальными областями: специфически между 7+ и 8-, больше число клеток-предшественников обнаруживалось в правой, чем в левой области . Эндотелиальные клетки предшественники не видны вне сердце-формирующих областей вплоть до стадии 8- (three somites). Свободные, мезенхимно-подобные клетки эндотелиальных предшественников являются внутренне присущими кардиальным областям. Они экспрессируют кроме того два внеклеточных антигена, JB3, a fibrillin-like protein, и cytotactin, оба ассоциированы с сегментами первичной сердечной трубки, где эндотелиальные клетки "re-transform" обратно в мезенхимные во время образования эндокардиальных подушек . Между стадиями 8 и 9 (4-7 сосмитов), (1) QH-1 позитивные клетки внутри сердце-формирующих областей образуют сосудисто-подобные ( vascular-like) соединения с QH-1 позитивными клетками, расположенными вне кардиальной области, (2) после слияния кардиальных областей ,образуется сплетение QH-1 позитивных клеток вентральнее передней кишки и (3) дефинитивная эндокардиальная выстилка первичной сердечной трубки образуется непосредственно из вентрального сплетения эндокардиальных клеток предшественников. Так как QH-1 позитивные эндотелиальные клетки предшественники каждой сердце-формирующей области находятся в тесной взаимосвязи с передней энтодермой , была предпринята попытка определить способствует ли энтодерма формированию клеток предшественников, предназначенных (committed) для кардиального эндотелиального клона по их экспрессии QH-1, JB3 antigen, и cytotactin (Sugi, Markwald, 1996). Для этого прекардиальные мезодермальные эксплантанты, взятые на стадии 5 из сердц-формирующих областей до экспрессии этими клетками эндокардиальных или миокардиальных маркеров культивировали на поверхности коллагена в присутствии или в отсуствии энтодермы. В отсуствие энтодермы прекардиальная мезодерма эксплантантов оставалась эпителиальной, формировала сократительную ткань и не давала QH-1 позитивных клеток или мезенхимных клеток . Энтодерма или ею обусловленная среда приводили к формировнию помимо контрактильной ткани к образованию мезенхимных клеток. Последние проникали в гель и как и in vivo экспрессировали QH-1 antigen, JB3 antigen, и cytotactin. Следовательно, энтодерма индуцирует в кардиальных полях эпителиально-мезенхимную трансформацию в результате которой происходит разделение миокардиальных и эндокардиальных предшественников.
Микроусловия мезодерм/энтодермального интерфейса пермиссивны и возможно инструктивны для эндотелиальной-эндокардиальной дифференцировки. Ряд молекул (фибронектин, факторы роста,включая FGF ,TGF-β семейства), секретируемых энтодермой способствует такой дифференцировке. Одними из самых ранних маркеров эндотелиальных клеток являются продукты генов Нех и flt-1 Эндокардиальные клетки сначала экспрессируют N-кадхерин, но затем начинают экспрессировать васкулярный кадхерин и отсортировываются от популяции кардиомиоцитов тем самым.
Имеются два независимых регуляторных пути: (1)один связан с развитием формы, трехмерного органа; (2) с регуляцией миогенеза. Воздействие оуабайна на очень ранней стадии ингибирует компартментализацию, формирование трехмерных эндокардиальных трубок и миогенез. Позднее (возможно во время эпителизации) тубулярная структура образуется, но миогенез остается подавленным. Затем, когда миогенный путь уже активирован, то оуабайн не способен проявлять свое ингибирующее влияние(Linask,Gui, 1995).