ГЛОМЕРУЛЫ ОБОНЯТЕЛЬНЫХ ЛУКОВИЦ
Зрелые обонятельные гломерулы являются многоклеточными ансамблями, где аксоны периферических обонятельных рецепторных нейронов (ОРН) синапсируют с дендритами вторичных проэкционных нейронов и интернейронов.
Гломерулы состоят из "оболочки" юкстагломерулярных (ЮГ) нейронов (проежуточных нейронов) и клеточных тел астроцитов, которые окружают глобулярный ансамбль нейропилей. Эти нейропили содержат синаптические контакты между аксонами ОРН, апикальными дендритами митральных и tufted клеток и дендритами ЮГ нейронов. Очень немногие тела клеток, первичная глия, локализованы в сердцевине гломерул. Отростки индивидуальных астроцитов ограничены исключительно одиночной гломерулой.
Предполагается, что гломерулы являются фукциональыми единицами инициальной стадии процессинга информаци о запахах. Имеются указания на то, что нейроны, экспрессирующие одни и те же одорантные рецепторыные гены конвергируют и синапсируют с двумя или немногими топографически фиксированными гломерулами луковицы.
У трансгенных мышей, у которых Р2 одорантный рецептор замещен Р3 одорантным рецептором, ОРН, экспрессирующие новый рецепторный ген проэцируются в гломерулы, соседние с гломерулами, предназначенными для ОРН с Р3 локусом. Следовательно, одорантные рецепторные гены участвуют в наведении аксонов или их активность необходима для коректного таргетинга. Нейрональная активность не только детерминирует инициальное формирование гломерул, но и может играть роль в детерминации их зрелой синаптической архитектуры. Аксон-глиальные гломерулы формируются еще в отсуствие ЮГ интернейронов. Кроме того гломерулы формируются нормально у мышей с отсуствием ЮГ интернейронов. Гломерулы формруются нормально у трансгенных мышей и в отсуствии митральныхю tufted клеток.
Индукция гломерул:аксон-глиальные взаимодействия.
Исследования (Bailey et al., 1999) показали, что перыми клеточными компонентами, образующими распознаваемые гломерул, являются радиальная глия/радиальные астроциты и аксоны ОРН.
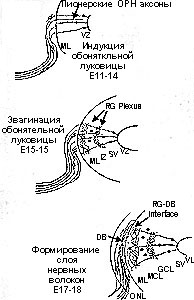
Рис. 1. Схема образования гломерул. Индукция обонятельных луковиц Е11-14:Достижение ОРН аксонами и сопровождающими обонятельными Шванновскими клетками рострального телэнцефалона. Часть аксонов проходит через мантийный слой (ML) и проникает в глубину вентрикулярного слоя (VL), большинство же ОРН аксонов остается вне телэнцефалона. Эвагинация обонятельных луковиц Е15-16: В результате изменений клеточных циклов в VL телэнцефалон эвагинирует. Формируются два новых слоя, субвентрикулярная зона (SV) и промежуточная зона (IZ). В это же время отростки радиальной глии (RG) луковицы формируют два сплетения, по сплетению с обеих сторон IZ. Формирование слоя нервных волокон Е17-18: ОРН аксоны энкапсулируют большую часть обонятельной луковицы в виде слоя нервных волокон (ONL) Во внутренней части этого слоя ОРН аксоны ранжированы в плотный диск (DB). Терминальные отростки RG формируют серии боковых веточек, непосредственно противопоставленные аксонам плотного диска, презумптивный "барьер". Инициация гломерул Е19-20: В ростральной части луковиц ОРН аксоны и отростки RG смешиваются и формируют протогломерулы, состоящие из аксональных гломерул и глиальных гломерул. Большинство гломерул содержит как глиальные, так и аксогнальные компоненты. Дендриты митральных клеток образуют обширное сплетение в средних областях луковиц, но не вносят вклада в гломерулы. Вовлечение JG клеток Е21-Р1: При рождении перикарии JG клеток мигрируют к периферии гломерул, формируюя перигломерулярные кольца из тел клеток. Дендриты митральных клеток присутствуют в гломерулярном слое, но еще не сформировали пучков (tufts) внутри гломерул (дендритных гломерул). Гломерулизация МС клеток Р4-Р5: Апикальные дендриты митральных клеток ограничены одиночными гломерулами. Эти апикальны отростки формируют четкие гломерулярные пучки (tufts) или дендритные гломерулы внутри отдельных гломерул. МС- митральные клетки, MCL - слой митральных клеток, GCL - слой гранулярных клеток, GL - слой гломерул, EPL - наружный плексиформный слой, IPL - внутренний плексиформный слой.
Стадии образования Обонятельных Луковиц (ОЛ)

(1) Индукция ОЛ. Е11-14.
Первые аксоны из обонятельного эпителия достигают ростральной части телэнцефалона. Субпопуляуия этих аксонов проникает через вентрикулярный слой и связана с изменениями в кинетике клеточных циклов в вентрикулярном слое
(2) Индукция ОЛ. Е15-16.
ОРН аксоны присутствуют в виде диффузной массы, форимурующей шапочку вокруг ростральной ОЛ, которая формируюется из эвагината теленэцефалического пузыря. Субпопцляция ОРН аксонов все еще глуоко проникает в вентрикулярную зону ОЛ, тогда как основаня масса аксонов остьается вне ОЛ. Радиальная Глия (RG) формируют сплетения поверхностнее промежуточной зоны и глубже этой зоны.
(3) Слой нервного волокна. Е17-18.
Количество ОРН аксонов на поверхности ОЛ увеличивается; эти аксоны формируют слой нервных волокон, окружающий большую часть ОЛ. ОРН аксоны в самой глубокой части слоя нервных волокон формируют плотный диск аксоновю Непосредствено рядом с ним серия латеральных RG отросков формирует "барьер" для большинства ОРН аксонов.
(4)Инициация гломерул. Е19-20.
В ростральной части плотного диска ОРН аксоны начинают коалесцировать в аксональные гломерулы. Одновеременно отростки RG формируют небольшие пучки или глиальные гломерулы. На ст. Е19-20 большинство протогломерул содержит как аксональные, так и глиальны компоненты, но некоторые состоят только из аксонов или глии.
(5)Вовлечение ЮГ клеток. Е21-Р1.
При рождении ЮГ клетки мигрируют в окружение гломерул в ростаральной части Ол, а дендриты митральных клеток все еще диффузно распределены.
(6)Гломерулизация дендритов. Р4-5.
Апикальные дендриты митральных клеток становятся ограниченными одиночными гломерулами. Апикальные отростки формируют пучки, которые располагаются внутри одиночной гломерулы.
(1) Способна ли глия рекрутировать аксоны в гломерулы.
Это м.б. связано с ингибирующим влиянием тенасцина или сульфатированных протеогликанов, экспрессируемых RG, на рост ОРН аксонов. Но глия может и способствовать росту аксонов. Часто она наводит аксоны к их мишеням.
(2) Способны ли аксоны рекрутировать глию в гломерулы.
Известно, что после удаления ОЛ-регенерирующие ОРН аксоны проникают в эктопические места ЦНС и формируют гломеруло-подобные структуры. Следовательно, формирование fasciculated balls of neuropil является внутренним свойством ОРН аксонов. Это подтверждается и экспериментами на моли.
(3) Аксоны и глия одновременно индуцируют гломерулы.
(4) Аксоны и Глия отвечают на общий внешний сигнал.
Juxtaglomerular (ЮГ) клетки и дендриты соединяют гломерулы.
Это м.б. связано с ингибирующим влиянием тенасцина или сульфатированных протеогликанов, экспрессируемых RG, на рост ОРН аксонов. Но глия может и способствовать росту аксонов. Часто она наводит аксоны к их мишеням.
ЮГ клетки начинают формировать кольца перикария, окружающие гломерулы на ст. Р0 в ростральной части и на стю З4-5 в каудальной части ОЛ. Значит эти клетки не участвуют в инициальных ступенях формирования гломерул, но участвуют в их стабилизации.
Было предположено, что обонятельные Шванновские клетки (периферическая глия), которые мигрируют в гломерулярный слой действуют, дробя гломерулярный нейропиль на протогломерулы. У моли она важна для стабилизации гломерулярной архетектуры. Остается неясным участвуют ли обонятельные Шванновские клетки у млекопитающих в индукции, стабилизации или других аспектах развития гломерул.
Обонятельные Швановские клетки, мигрирующие из обонятельного эпителия в ONL экспрессируют высокие уровни рецепторов факторов роста нерввов низкого сроства (LNGFr) и S100. Однако эти клетки никогда не проникают в глубокие части ONL. ЭМ исследования показали, что обонятельные Шванновские клетки присутствуют в iONL и в гломерулярном слое. Отсутствие окрашивания LNGFrюS100 в этих обонятельных Шванновских клетках, по-видимому, обусловлено задержкой/градиентом созревания, но возможно что эти клетки предстваляют собой временно специализированную глию, которая позднее трансформируется в типичные обонятельные шванновские клетки.
Однако в другой работе ( Valverde, 1999)высказывается несколько иная точка на роль обонятельных Шванновских клеток. Предполагается, что оболочечная глия наводит ОРН аксоны
Обонятельные аксоны начинают выходить из обонятельной плакоды около ст. Е12. Рост аксонов всегда сопровождается клетками презумптивной оболочечной глии, также происходящих из обонятельных плакод. Эти клетки сохраняют свои ассоциации до тех пор, пока они не установят контакты с зачатком ОЛ. Многочисленные митотические фигуры обнаруживаются в популяции сопровождающих клеток во время этого онтогенетического процесса. Эта пролиферация может объяснить результирующую большую массу клеток (миграторную массу), достигающих ОЛ на поздних стадиях. Кроме того затянувшаяся незрелость оболочечных клеток может облегчить сортировку обонятельных аксонов в соответстви с их одор-специфичностью до формирования спцифических гломерул.
Первые синаптические контакты обнаруживаются на Е18 в области, соответствующей территории, окупируемой миграторной массой. Синаптические окончания выглядят более сконцентированными в дискретных локусах плотных нейропиль содержащих дендритных отростков митральных/туфтовых клеток и синаптические окончания окружены отростками оболочечных клеток, которые формруют стенки, отделяющие эти зоны высокой синаптической плотности. Предполагается, что эти функциональные (синаптические) домены представляют собой инициальное формирование обонятельных гломерул, описанное Baley и соавт.
Автор подчеркивает важность оболочечной глии (обонятельных Шванновских клеток) в наведении аксонов в направлении их предназанчения. Оболочечная глия обеспечивает необходимые механизмы для выбора аксонами ОРН пути к удаленным гломерулам.
Рассматривая стадии инициации гломерул Baily с соавт. отрицают возможность участия в формировании гломерул других компонентов кроме аксонов ОРН и отростков RG. (Treolar et al., 1999) показали дендриты митральных/туфтовых клеток формируют "дендритную зону" (которая описывается и Baleys...) непосредственно противостоящую слою нервных волокон, формируемых аксонами ОРН. Они полагают, что ОРН аксоны индуцируют формирование гломерул около Е19 в результате врастания в эту дендритную зону. Если Bailey с соав. делают упор на образовании глиальных гломерул, то Treloar с соавт. подробно рассматривают пространственно-временные последовательности формирования синапсов на тканевых срезах с двойным мечением, которые,по-видимому, происходят чуть раньше, чем формирование глиальных гломерул.