ИННЕРВАЦИЯ КОРТИЕВА ОРГАНА
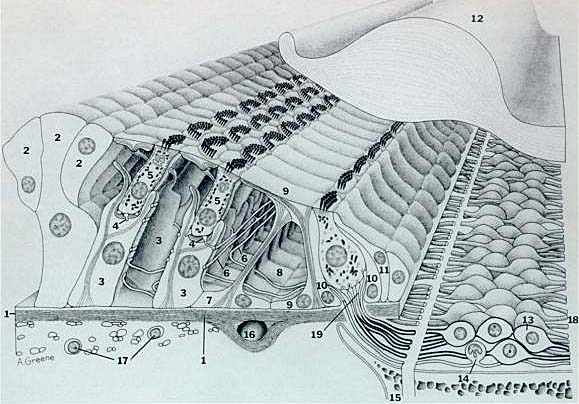
Разрез типичного базального витка улитки млекопитающих. Диаметр наружной волосковой клетки примерно 7 µm. Пустые пространства в основаниях наружных волосковых клеток заняты эфферентными окончаниями, которые не видны на рис. 1 - базилярная мембрана, 2 - Hensen клетки, 3 - наружные фалангеальные клетки Deiters, 4 - нервные окончания, 5 - наружные волосковые клетки, 6 - наружные спиральные волокна, 7 - наружные pillar клетки, 8 - внутренний туннель, 9 - внутриенние pillar клетки, 10 - внутренние фалангиальные клетки, 11 - пограничные клетки, 12 - текториальная мембрана, 13 - клетка спирального ганглия типа I, 14 - клетка спираьного ганглия типа II, 15 - костная спиральная ламина, 16 - спиральный кровеносный сосуд, 17 - веретенообразные клетки, 18 - аксоны клетки спирального ганглия (волокна слухового нерва), 19 - радиальные волокна.
Оливо-кохлеарная система поставляет эфферентную иннервацию в орган Корти.
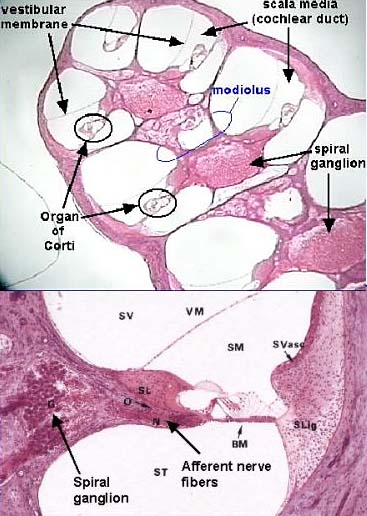
Вызванные звуком колебания базилярной мембраны возбуждают высвобождение нейротрансмиттера из внутренних волосковых клеток в отростки биполярных нейронов чьи клеточные тела расположены в спиральном ганглии. Этот ганглий находится внутри улитки, рядом с ее костной центральной осью, modiolus. Аксоны от нейронов спирального ганглия проецируются по нерву VIII в ствол мозга.
Метили кохлеарные волокна от latheral superior olivary (LSO) ядра и билатерально от ventral nucleus trapezoid body (VNTB)(рис.1)
У новорожденных крыс (Р0) меченные оливокохлеарные эфферентные аксоны приближаются к внутренним волосковым клеткам и к наружным на 2-й день постнатального развития (Р2). Незрелые эфферентные аксоны обнаруживаются среди поддерживающих клеток большого эпителиального гребня, а также между внутренними волосковыми клетками (Р0). Волосковые клетки, первыми вступающие в контакт, содержат небольшое число нерегулярного размера пузырьков и митохондрий. Постсинаптические специализации во внутренних волосковых клетках содержат субсинаптические цистерны, нерегулярного размера пузырьки и синаптические тела. Сходные структуры образуются с окончаниями афферентных волокон. Эфферентные волокна вступают в синапс с наружными волосковыми клетками на ст. Р4. Контакты между афферентными и эфферентными волокнами часто обнаруживаются во внутреннем спиральном пучке со ст. Р6. По мере созревания окончаний эфферентных волокон все больше синаптических пузырьков и постсинаптических цистерн появляется в волосковых клетках. С Р9 по Р11 волосковые клетки могут реагировать на внешние слуховые стимулы, однако афферентные и эфферентные волокна не могут.
Полученные результаты показывают 1) эфферентные аксоны контакируют с IHCs и образуют незрелые контакты на ст. Р04; 2) эфферентные аксоны растут вдоль наружных pillar клеток и контактируют с OHCs на ст. Р2 и образуют с ними синапсы на ст. Р4. и 3) эфферентные аксоны образуют синапс-подобные контакты с Deiters' клетками на ст. Р4.
Факторы наведения эфферентных волокон
BDNF и NT-3 и их специфические с высоком сродством тирозин киназыне рецепторы , trkB и trkC, обязательны для выживания кохлеарных афферентных нейронов и для наведения их в направлении органа Корти. Эфферентные волокна, по-видимому, слудуют молекулярным сигналам, издаваемыми афферентными волокнами, чтобы достигнуть органа Корти, так как эфферентные волокна неспособны достигать органа Корти у мутантных мышей, у которых отсутствуют афферентные волокна, хотя нейротрофины не имзенены.
;По достижении эфферентными волокнами волосковых клеток они м. реагировать на локальные сигналы, чтобы найти соотв. синаптические мишени. Гликопротеины ВКМ, такие как тенасцин-С, м. управлять локальным афферентным и эфферентным ростом. Взаимодействия между нейротрансмиттерами, высвобождаемыми эфферентными окончаниями и их рецепторами на волосковых клетках и афферентных дендритах м. играть роль в формировании синапсов. Предполагается, что появление α9 nicotinic acetyulcholine receptor subunits в волосковых совпадает с эфферентной иннервацией и м. влиять на наведение эфферентных волокон. Однако согласно другим данным экспрессия этой рецепторной субъединицы отражает состояние внутреннего созревания волосковых клеток и не зависит от эфферентных волокон. Предполагается, что BDNF и trkB являются кандидатами, обеспечивающими локальную эфферентную реогранизацию, т.к. пространственные и временные изменения их экспресии совпадают с изменениями в эфферентной иннервации. BDNF локализуется в волосковых, поддерживающих клетках и клетках спирального ганглия в виде паттерна, меняющегося в пространстве и времени. Однако эфферентные влокна не продуцируют еклИ полной длины, принципиального рецептора для BDNF.
NT-3 и trkC также являются кандидатами на роль модуляторов локальной эфферентной специфичности в органе Корти. Эфферентные волокна слабо позитивны по trkC рецепторам, а NT-3 присутствует в поддерживающих клетках Кортиева органа, включая большой эпителиальный гребень и Deitetrs' клетки.
Эфферентный синаптогенез в IHCs
Афферентные волокна достигают органа Корти раньше эфферентных.Эфферентные волокна контактирующие с основанием и сторонами ШРСы на ст. Р0 слабо дифференцированы и содержат немного нерегулярного размера пузыьков. Различные специализации в IHCs напротив этих аксонов содержат цистерны, аггрегаты большим и малых пузырьков и по-видимому, синаптичские тельца.
Синапс-подобные контакты с Deiters' клетками
Синапс-подобные контакты между эфферентными волокнами и Вушеукыэ клеткаи наблюдаютсяна ст. Р4-6. У молодых животных эти клетки секретируют компоненты минорной текториальной мембраны вмсте с pillar и Гензеновскими клетками. Эфферентные контакты могут модулировать секрецию этих клеток. Дейтеровкие клетки, по-видимому, нарушают кохлеарную мехаику в ответ на эндогенный АТФ как и на звук и эфферентные контакты могут модулировать эту реакцию. В это время эфферентные аксоны кажутся спиральными, проходя между рядами Дейтеровских клеток вдоль базилярной мембраны. Кохлеарные афферентные волокна типа II контактируют с Дейтеровскими клетками у взрослых кошек и морских свинок, но синаптических контактов не обнаружено.