введение
Дорсо-вентральная и передне-задняя оси, а следовательно, и билатеральная симметрия выявляются во время гаструляции. Морфологически обнаружимая асимметрия обнаруживается в позднем эмбриогенезе после образования петли сердечной трубки и киши [1] [2].
Образование кародиальной петли высоко законсервировано у позвоночных [3]. Обнаружены асимметрично экспрессирующиеся гены у эмбрионов кур [4], более 30 мутантных генов, затрагивающих лево-правостороннее развитие, открыто у позвоновных (reviewed in [5•]). Результаты пересадок ткани указывают на то, что лево-правосторонняя ориентация специфицируется задолго до образования петель. Ключевые гены, которые функционируют позже, такие как pitx2 и cSNR1, являются транскрипционными факторами, экспрессирующимися в left–right органных клонах.
Conservation of asymmetric expression patterns in lateral plate mesoderm
Nodal, член семейства TGF-β
межклеточных сигнальных молекул,
экспрессируется в левой латеральной
пластинке мезодермы кур [4],
мышей [6] [7],
лягушек [7] [8]
и рыбок данио [9•] [10•].
Большая часть мезодермы латеральной
пластинки, в которой экспрессируется nodal,
вносит вклад в билатеральрно симметричные
структуры, однако соседняя мезодерма дает
асимметричные структуры. Паттерны
экспрессии nodal нарушаются у laterality mutants мышей,
таких как iv [7], inv [6] [7], ft [11]
и no turning [12]. IV
кодирует left–right dynein (LRD) [13],
а INV кодирует inversin, внутриклеточный
белок с Ank/Swi6 повторами [14•] [15•].
кроме того, экспрессия nodal нарушается при
действии на ранних эмбрионов факторов,
нарушающих laterality, таких как нарушение
экспрессии Sonic hedgehog (Shh), activin или FGF8 у кур [4] [16••] [17],
экспрессии Vg1 или семейства доминантно
негативных рецепторов TGF- β
у Xenopus [18] [19••],
экстирпация клеток средней линии [8] или нарушения щелевых соеднинений [20••].
анализ эктопической экспрессии nodal на
правой стороне у кур [21]
или лягушек [22] выявил
стохастичность ориентации сердца. т.о., nodal
скорее всего выступает как медиатор между ранними сигналами и
спецификацией left–right информации в
зачатках сердца и др. органов.
Lefty2, другой член семейства TGF-β , также экспрессируется в левой латеральной пластинке мезодермы мыши [23] [24]. как и в случае nodal, паттерн экспрессии Lefty2 нарушен у laterality мутантов iv [23], inv [23], ft [11], nt [12], lefty1 [25••] and KIF3B [26••]. семейство Lefty является важным компонентом left–right пути [25••] [27•] [28•].
Downstream of Nodal: Pitx2 and the execution of left–right morphogenesis
Pitx2, член семейства bicoid гомеобокс-содержащих транскрипционных факторов отвечает за синдром Reiger [29] и экспрессируется билатерально в цефалической мезодерме и ткани гипофиза. Pitx2 экспрессируется на левой стороне кардиальной трубки и кишки у кур [30•] [31••] [32••] [33••] [34•] [35••], мышей [[25••], [32••] [33••] [34•]], frog [33••] [35••], и рыбок данио [28•] [35••]. асимметричная экспрессия pitx2 персистирует в зачатках органов после временной асимметричной экспрессии nodal .
две линии доказательств указывают на то, что pitx2 является нижестоящим компонентом на left–right signaling пути. во-первых, паттерн экспрессии pitx2 нарушается у laterality мутантных мышей, коррелируя с нарушением ориентации органа. Гомозиготные iv/iv эмбрионы имеют стохастическую ориентацию органов и паттерн экспрессии (билатеральный, левый, правый или отсутствие экспрессии) pitx2 [32••] [33••] [35••]. Передний домен экспрессии pitx2 является билатеральным у мутантов lefty1, которые имеют торакальный левый изомеризм [25••]. У inv/inv эмбрионов left–right ориентация органов инвертирована (зеркальна по отношению к норме) и pitx2 экспрессируется на правой, а не на левой стороне [33••]. Во-вторых, паттерн экспрессии pitx2 у кур или Xenopus нарушается эктопической экспрессией left–right генов (reviewed in [5•]). у кур, эктопическая экспрессия Shh справа индуцирует билатеральную экспрессию pitx2 [30•] [31••] [32••] [33••]. Shh-блокирующие антитела слева уменьшают слева нормальную экспрессию left-sided nodal [16••] и pitx2 [31••]. эктопическая экспрессия nodal справа индуцирует pitx2 на правой стороне у эмбрионов кур [31••] [32••] [33••] или лягушек [35••]. следовательно, nodal стоит выше pitx2 , а Shh , по крайней мере у кур, выше обоих.
эктопическая экспрессия pitx2 у кур [31••] [33••] или лягушек [35••] нарушает left–right развитие и оказывает дополнительные эффекты. эктопическая экспрессия pitx2 справа не индуцирует nodal и не нарушает экспрессии cSNR1 [31••], который обычно экспрессируется в правой латеральной пластинке мезодермы [36]. эктопическая экспрессия pitx2 у кур увеличивает частоту кардиального изомеризма [31••] [33••]. Logan et al. [31••] предприняли попытку одновременно элиминировать Shh–Nodal–Pitx2 путь слева с помощью Shh-блокирующих антител и активровать pitx2 справа с помощью инфекции pitx2-экспрессирующим вирусом. это устраняло кардиальный изомеризм и давало left–right инверсию у 30% эмбрионов.
таким образом, pitx2 left–right асимметричная экспрессия высоко законсервирована у позвоночных и pitx2 находится ниже nodal. должна быть батарея генов, которые регулируются с помощью pitx2 и ко-факторов во время left–right морфогенеза.
Upstream of Nodal: left–right coordinator, node and midline
три отдельных сигнальных центра должны обеспечивать на паттерн асимметричной экспрессии nodal: клетки латеральнее узла, называемые left–right coordinator (LRC); клетки узла, посредством асимметричной экспрессии генов и/или monocilia activity; и клетки средней линии. экспериментны на эмбрионах лягушек подтвердили, что асимметричный процессинг белка члена семейства TGF-β , Vg1, на левой латеральной стороне эмбрионов лягушки иницирует лево-правостороннюю ось [19••]. эктопическая экспрессия Vg1 после процессинга на правой стороне инвертирует лево-правостороннюю ось, включая экспрессию nodal [19••]. соответственно, экспрессия доминантно-негативных рецепторов или лигандов, которые взаимодействуют с Vg1 signaling на левой стороне эмбриона ведет к стохастическому left–right развитию [19••]. хирургическая ротация узелка у ранних эмбрионов кур указывает на то, что возникновение ассиметричной экспрессии генов в узелке зависит от сигналов, возникающих вне узелка [16••], вообще-то латеральные клетки сравнимы с LRC лягушек. осложняется еще тем, что асимметричная экспрессия в узелке отсутствует у non-avian эмбрионов.
Клетки узелка имеют специализированные monocilia [26••] [37] [38] , которые дефектны у мутантов по kinesin KIF3B [26••], kinesin KIF3A [39•] or hepatocyte nuclear factor/forkhead homologue 4 [40•]. LRD также экспрессируется в клетках узелка [13]. соответственно, каждый из этих мутантов имеет laterality дефекты. Движения узелковых моноцилий (monocilia) могут управлять асимметричным распределением внеклеточных частиц, они дефектны у KIF3B мутантов [26••] (reviewed in [41]). Однако вряд ли асимметричные движения monocilia формируют барьеры между двумя сторонами узелка или окружающей латеральной тканью, препятсвуя дисперсии информации (Fig. 1). Клетки, которые мигрируют через узелок во время гаструляции образуют срединную линию эмбриона, хорду, hypochord и вентральную донную пластинку нервной трубки. Средняя линия является существенной для left–right развития или путем формирования барьера или путем передачи сигналов латеральным тканям [5•] [42] [43].
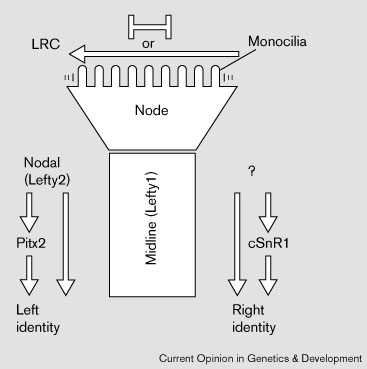 |
|
Figure 1 Left–right pathway. Left–Right Coordinator (LRC) и узелок участвуют в инициации left–right развития. У мышей, моноцилии узелка движут внеклеточные частицы влево. Движения моноцилий обеспечивают перераспределние сигнальных молекул , которые эксспресируются симметрично в латеральной ткани (стрелка влево), или создают барьер (bar signs)для поддержания различий между двумя сторонами узелка, которые инициируются в латеральной ткани (LRC). Некоторые гены экспрессируются асимметрично внутри узелка кур. Эти ранние события обеспечивают асимметричную экспрессию nodal и lefty2 слева (у кур) cSnR1 справа. Поддержание асимметрии в латеральной пластинке мезодермы зависит от эмбриональной срединной линии [5•]. |
Conclusions and perspectives: diverse origins that converge at left–right phylotypic stages?
Модель с привлечением LRC, узелка и средней линии не является, по-видимому, исключительной (Fig. 1). Столь же вероятно, что разные группы позвоночных используют разные механизмы инициации left–right оси и каждый механизм конвергирует с асимметричным распределением nodal, lefty и pitx2 (Fig. 2).
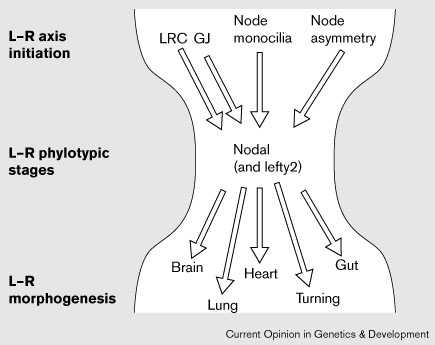 |
Figure 2 Model: the left–right (L–R) phylotypic stages. The Left–Right Coordinator (LRC), gap junction communication (GJ), node monocilia movement and asymmetric gene expression in the node have each been implicated in the initiation of left–right development and the establishment of nodal expression on the left side of the embryo. The mechanisms that initiate left–right axis formation might have diverged in an individual taxon; however, each passes through a constrictive phase of highly conserved nodal expression patterns, during the left–right phylotypic stages, analogous to the phylotypic 'developmental hourglass' stages indicative of the zootype along the anterior–posterior axis (reviewed in [45]). Although the pathway to left–right morphogenesis can be perturbed late in development, the initiation of the left–right axis occurs early, either before or coincident with the formation of the node at the beginning of gastrulation, and is specified in lateral tissues during early neurula stages. It is striking that the proposed left–right phylotypic stages are coincident with the phylotypic stages of Hox gene patterning along the anterior–posterior axis. Each individual organ primordium derives its left–right information from the phylotypic stages, which subsequently regulates unique aspects of organ morphogenesis. |
Я предполагаю, что имеются 'left–right zootype' , определяемые с помощью законсервированного паттерна left–right экспрессии генов (Fig. 2). Такой 'left–right zootype' в чем-то анологичен anterior–posterior zootype, предложенному Slack et al. [44], который базируется на паттерне законсервированной экспрессии HOX генов (reviewed in [45]); однако, left–right zootype м.б. ограничен хордовыми.
В модели left–right zootype стадии высоко законсервированной nodal (и вообще-то lefty2 и pitx2) экспрессии в левых латеральных тканях отражают спецификацию лево-правосторонней оси и м.б. определены как 'left–right phylotypic stages' для индивидуального таксона (Fig. 2). Это указывает на то, что по крайней мере одна чать пути вниз от nodal, и вообще кардиальный органогенез, высоко чувствительны к селективному давлению , дающему в результате асимметричную экспрессию nodal. Ступени на пути к верхним left–right phylotypic stages должны быть способными к дивергенции. Позднее в ходе развития, после left–right phylotypic stages, генетические пути, которые регулируют морфогенез left–right асимметрии различных органов , таких как сердце, кишка и головной мозг могут дивергировать и даже теряться для некоторых структур в разных таксонах.
литература
Linkage of cardiac left-right asymmetry and dorsal-anterior development in Xenopus.
Development
1995,
121:
1467–1474.
Vertebrate left-right development.
Cell
1995,
82:
689–692.
Cardiac design in lower vertebrates: what can phylogeny reveal about ontogeny?
Experientia
1988,
44:
919–930.
A molecular pathway determining left-right asymmetry in chick embryogenesis.
Cell
1995,
82:
803–814.
Molecular mechanisms of vertebrate left-right development.
Trends Genet
1998,
14:
459–465.
Relationship between asymmetric nodal expression and the direction of embryonic turning.
Nature
1996,
381:
155–158.
Conserved left-right asymmetry of nodal expression and alterations in murine situs inversus.
Nature
1996,
381:
158–161.
Left-right asymmetry of a nodal-related gene is regulated by dorsoanterior midline structures during Xenopus development.
Development
1997,
124:
1465–1472.
Induction of the zebrafish ventral brain and floorplate requires cyclops/nodal signalling.
Nature
1998,
395:
185–189.
Zebrafish nodal-related genes are implicated in axial patterning and establishing left-right asymmetry.
Dev Biol
1998,
199:
261–272.
The expression pattern of nodal and lefty in the mouse mutant Ft suggests a function in the establishment of handedness.
Mech Dev
1997,
66:
5–11.
No turning, a mouse mutation causing left-right and axial patterning defects.
Dev Biol
1998,
193:
77–89.
Mutation of an axonemal dynein affects left-right asymmetry in inversus viscerum mice.
Nature
1997,
389:
963–966.
Cloning of inv, a gene that controls left/right asymmetry and kidney development.
Nature
1998,
395:
177–181.
Inversin, a novel gene in the vertebrate left-right axis pathway, is partially deleted in the inv mouse.
Nat Genet
1988,
20:
149–156.
The transfer of left-right positional information during chick embryogenesis.
Cell
1998,
93:
25–35.
FGF8 functions in the specification of the right body side of the chick.
Curr Biol
1999,
9:
277–280.
Initiation of vertebrate left-right axis formation by maternal vg1.
Nature
1996,
384:
62–65.
The left-right coordinator: the role of Vg1 in organizing left-right axis formation.
Cell
1998,
93:
37–46.
Gap junctions are involved in the early generation of left-right asymmetry.
Dev Biol
1998,
203:
90–105.
Left/right patterning signals and the independent regulation of different aspects of situs in the chick embryo.
Dev Biol
1997,
189:
57–67.
Functional differences among Xenopus nodal-related genes in left-right axis determination.
Development
1997,
124:
3293–3302.
Left-right asymmetric expression of the TGF-β family member lefty in mouse embryos.
Nature
1996,
381:
151–155.
Two closely-related left-right asymmetrically expressed genes, lefty-1 and lefty-2: their distinct expression domains, chromosomal linkage and direct neuralizing activity in Xenopus embryos.
Genes Cells
1997,
2:
513–524.
lefty-1 is required for left-right determination as a regulator of lefty-2 and nodal.
Cell
1998,
94:
287–297.
Randomization of left-right asymmetry due to loss of nodal cilia generating leftward flow of extraembryonic fluid in mice lacking KIF3B motor protein.
Cell
1998,
95:
829–837.
Antivin, a novel and divergent member of the TGFbeta superfamily, negatively regulates mesoderm induction.
Development
1999,
126:
229–240.
Regulation of midline development by antagonism of lefty and nodal signaling.
Development
1999,
126:
3253–3262.
Cloning and characterization of a novel bicoid-related homeobox transcription factor gene, RIEG, involved in Rieger syndrome.
Nat Genet
1996,
14:
392–399.
Cloning and expression pattern of chicken Pitx2: a new component in the SHH signaling pathway controlling embryonic heart looping.
Biochem Biophys Res Commun
1998,
247:
100–105.
The transcription factor Pitx2 mediates situs-specific morphogenesis in response to left-right asymmetric signals.
Cell
1998,
94:
307–317.
Pitx2 participates in the late phase of the pathway controlling left-right asymmetry.
Cell
1998,
94:
319–324.
Pitx2 determines left-right asymmetry of internal organs in vertebrates.
Nature
1998,
394:
545–551
Pitx2, a bicoid-type homeobox gene, is involved in a lefty-signaling pathway in determination of left-right asymmetry.
Cell
1998,
94:
299–305.
The homeobox gene Pitx2: mediator of asymmetric left-right signaling in vertebrate heart and gut looping.
Development
1999,
126:
1225–1234.
Control of vertebrate left-right asymmetry by a snail-related zinc finger gene.
Science
1997,
275:
1301–1304.
Morphogenesis of the murine node and notochordal plate.
Dev Dyn
1994,
201:
260–278.
Cell proliferation in mammalian gastrulation: the ventral node and notochord are relatively quiescent.
Dev Dyn
1996,
205:
471–485.
Situs inversus and embryonic ciliary morphogenesis defects in mouse mutants lacking the KIF3A subunit of kinesin-II.
Proc Natl Acad Sci USA
1999,
96:
5043–5048.
Mutation of the mouse hepatocyte nuclear factor/forkhead homologue 4 gene results in an absence of cilia and random left-right asymmetry.
J Clin Invest
1998,
102:
1077–1082.
A new spin on handed asymmetry.
Nature
1999,
397:
295,297–298.
Maintenance of asymmetric nodal expression in Xenopus laevis.
Dev Genet
1998,
23:
194–202.
Evolutionary conservation of mechanisms upstream of asymmetric Nodal expression: reconciling chick and Xenopus.
Dev Genet
1998,
23:
185–193.
The zootype and the phylotypic stage.
Nature
1993,
361:
490–492.
Sornite number and vertebrate evolution.
Development
1998,
125:
151–160.