В ходе раннего эмбрионального развития позвоночных происходят разнообразные процессы сегментации исходных осевых структур: формировние нейромеров, сомитомеров и сомитов, краниальных и спинальных ганглиев, нервов, бранхиальных дуг и т.д.(Мглинец, 1995)Сходные процессы сегментации нервной трубки происходят и у эмбрионов человека. Согласно нейромерной теории развития ЦНС задний мозг содержит 8 нейромеров, называемых ромбомерами (r1-r8), которые располагаются в рострокаудальной последовательности. Передний мозг содержит 6 нейромеров, называемых просомерами (р6-р1), которые располагаются в каудоростральной последовательности. Средний мозг представлен единственным нейромером.
|
Предполагается, что основу нейромеров составляют катехоламинэргические нейроны, выявляемые по их тирозин-гидроксилазной иммунореактивности (TH-IR) (Puels, Verney ,1998).
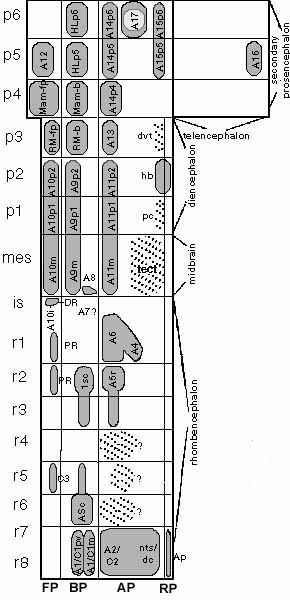
Рис.1. Топологическая модель продольных и нейромерных доменов головного мозга, иллюстрирующая относительное морфологическое положение различных тирозин-гидроксилаза иммунореактивных (TH-IR) популяций клеток. Продольные зоны обозначены внизу, нейромерные элементы - слева, а большие морфологические единицы обозначены справа. Кружок в р6 представляет собой глазной стебелек. Различные TH-IR (показаны серым - постоянные, точками - временные)области обнаруживают тенденцию располагаться вдоль определенных продольных зон и тем самым обнаруживают повторяющиеся паттерны в разных сегментах. Морфологические взаимотношения не отражают точных пропорций; тем не менее размеры разных областей отражают насыщенность их TH-IR клетками. Использована терминология Dahlstrom, Fuxe (1964) за некоторыми исключениями: А2/С2 термин зарезервирован за нейронами в ядрах тракта solitarius и дорсального столба ядер (алярные элементы каудальной части продолговатого мозга); группы клеток, развивабющиеся вокруг моторного столба vagus названы "перивентрикулярной частью А1/С1" (А1/С1pv), тогда как классический А1/С1 переименован в "мигрирующую часть А1/С1" (А1/С1m); группа А5 подразделена на независимые каудальную и ростральную части (А5с, А5r); периакведуктальная группа срелднего мозга переименована в "А11m". Классические группы среднего и переднего мозга (А9, А10, А11, А14, А15) обычно формируют многосегментные комплексы, чьи нейромерные части отличаются добавлением соответствующих суффиксов (i, m, p1-p6). Маммилярное и ретромаммлилярное образования обозначены в соответствии с их происхождением в базальной или или потолочной пластинке (суффикс -b или -fp). Группа клеток А7 , описанная у других млекопитающих, в данной работе не обнаружена; ее потенциальное топологическое положение обозначено А7?. Временные группы клеток r4-r6 алярной пластинки также обозначены ?. Некоторые популяции заднего мозга, по-видимому, родственны с областью шва (С3, PR, DR и А10i; А10i является поздно-появляющимся компонентом перешейка вентральной области тегментума) FP - floor plate, BP - basal plate, AP - alar plate, PR - pontine raphe Fig.1. Topological model of the longitudinal and neuromeric domains of the brain, illustrating the relative morphological position of the diverse tyrosine-hydroxylase (TH) immunoreativity (IR) populations. The longitudinal zones are indicated at the bottom, the neuromeric elements are indicated at the left side, and larger morphological units are identified at the right side. The circle in p6 drawn with a thicker line represents the eye stalk. The different TH-IR areas (gray areas indicate permanent populations, dashed areas transient ones) tend to line up along particular longitudinal zones and thus show a repeating pattern in diverse segments. Representation conserves only the mutual morphologically significant relationships and not true proportions; nevertheless, the sizes of the different areas depicted aim approximately to indicate te relative abundance of TH-IR cells. The terminology by Dahlstrom and Fuxe (1964) is followed in large measure, with some exceptions: the A2/C2 term is reserved for neurons within the nucleus tractus solitarius and dorsal column nuclei (alar elements of caudal medulla); the cell group developing around the vagal motor column is called the "periventricular part of A2/C1" (A1/C1pv), whereas the classical A1/C1 is reidd the "migrated part of the A1/C1 (A1/C1m); the A5 group is divided into independet caudal and rostral parts (A5c, A5r); and the midbrain periacueductal group is reidd "A11m". Classical groups of the midbrain and forebrain (A9, A10, A11, A14, A15) typically form plurisegmental complexes, whose neuromeric parts are distinguished by appending the corresponding suffix (i, m, p1-p6). Mammillary and retromammillary formations lack and alpha-numeric identification and are divided according to their origin in the basal plate or floor plate (suffix -b or -fp). The A7 cell group described in other mammals was not found in the present study; its potential topological position is indicated according to our tentative interpretation of the date in the literature (A7 ?). Transient cell groups of the r4-r6 alar plate are also labeled with interrogation signs because their significance is the unknown. Several hindbrain populations appear to be related to the raphe region (C3, PR, DR, and A10i; A10i is a late-appearing isthmic component of the ventral tegmental area). Вот доказательства сегментного паттерна распределения TH-IR групп клеток у самых ранних из изученных эмбрионов человека (стадия 15). Показано, что TH-IR нейроны возникают в виде специфических участков нейроэпителия, часто разделенных поперечными границами, совпадающими с межсегментными границами. Четкие границы выявлены (1) для примордиев tuberomammillary и supraoptoparaventricular primordia в р1, (2) для дорсальной (преходящей) и вентролатеральной (ZI) алярной группы клеток в р3, (3) для habenular популяции в р2, (4) для раннего рострального ограничения тектальных клеток на границе р1/mes, (5) для каудального конца nigral и ventral tegmental formations на границе mes/ist, (6) для каудального конца locus coeruleus complex (А6,А4) на r1/r2 границе, (7) для ранних дорсальных групп клеток относящихся к ромбомерам r2 и r4, (8) для парамедианных базальных групп клеток в r2и d r6 и (9) для рострального конца А1/С1-А2/С2 групп на границе r6/r7.
Помимо временно позитивных групп TH-IR популяции нейронов увеличиваются в течение наблюдаемого периода (36-44 день), однако они остаются топологически фиксированными в пределах тех же самых границ, независимо от существенных морфогенетических деформаций стенки головного мозга и радиальной миграции (например супраоптопаравентрикулярного комплекса). Предполагается, что процесс сегментации обусловливается генами, специфицирующими региональную идентичность (Мглинец, 1995), (Puels, Verney ,1998). Области нервной трубки способны продуцировать TH-IR клетки четко разграниченные в отношении продольных подразделений трубки на сегменты ( возможно и в отношении дорсо-вентрального паттерна). Это наблюдение приложимо в целом и в отношении экспрессии других молекул, таких как ацетилхолинэстераза, FORSE-1 бклок, BEN белок или кадхерины, маркеры ранней нейрональной дифференцировки. Четко разграниченный паттерн экспрессии обнаруживается и в отношении базальной и алярной пластинки.
TH-IR нейроны возникают независимо вдоль всей оси головного мозга человека, макакк резус. Очевидно, что индуктивные эффекты вентральной средней линии существенны для возникновения катехоламиновых групп клеток в парамедианных и базальных пластинках, и в меньшей степени для популяций алярных пластинок. Потеря функции вентрально экспрессирующегося гена Nurr-1 ведет к селективному дефициту вентрально орасположенных А9/А10 катехоламиновых групп клеток. Напротив близость ряда алярных TH-IR групп клеток к потолочной пластинке ( дорсальные алярные группы в р1-р3 и клетки области postrema в r7-r8), по-видимому, позволяет ассоциировать их с индуктивными сигналами, производными потолочной пластинки.
Большая часть TH-IR популяций каудальнее истмомезэнцефалической границы дифференцируется слс в норадренэргические или адренэргические нейроны, тогда как популяуция TH-IR в среднем и переднем мозге становится в основном допаминэргическими нейронами. Возможным фактором, участвующим в предопределении подобных различий, может быть Otx2 ген, именно его жомен экспресси заканчивается на перешейке.
Источник ростральных А9/А10 нейроны уже выявляется на 4,5 неделе развития до существенной миграции мантийных клеток. Это указывает на то, что А9/А10 комплекс возникает из нескольких сегментов (просомеров р1-р3) и среднего мозга. Сравнительная эмбриология ядра акцессорного оптического тракта указывает на то, что это ядро возникает в р1. Следовательно, часть А9/А10 комплекса до этого ядра возникает из диэнцефалона, а часть после этого ядра из мезэнцефалона.
На схеме представлена топологическая карта TH-IR групп клеток
Наиболее существенные перемещения TH-IR клеток наблюдаются в радиальных направлениях в базальной пластинке( в группах А1/С1m, A5c, A5r,A8,A9,HL) и в алярной пластинке ( группы спинальных дорсальных рогов, A2/C2, lsc, и А14 супраоптический компонент. Нейроны тегментальной области (А10) также мигрируют радиально внутри донной пластинки из глубокого положения, где они,по-видимому, смешаны с нейроэпителиальными клетками, в последующее subpial положение. Радиальную миграцию направляет радиальная глия.
Другим типом миграции (напр., комплекс А9) является достижение базальными клетками subpial положения. Эти клетки перемещаются тангенциально в латеродорсальном направлении с каждой стороны донной пластинки, которые вместе дают компактный слой субстранции нигра. Это перемещение может идти по пути следования их собственных аксонов.
Ассоциация мигрирующих TH-IR нейронов ствола мозга с мигрирующими двигательными нейронами может обеспечиваться независимым наведением вдоль одних и тех же путей. Но между ними может существовать и причинная связь или они могут отвечать на один и тот же сигнал. В этом случае двигательные нейроны должны сначала двигаться в направлении TH-IR клеток. Например, двигательные нейроны лицевого нерва из r4 идут сначала каудально проходя через ядро abducens в r5, а затем вентролатерально к дефинитивной поверхностной локализации в r6, где популяция TH-IR клеток значительно больше, чем в r3.
|