The nuclear pore complex: from molecular architecture to functional dynamics
Daniel Stoffler, Birthe Fahrenkrog, Ueli Aebi
Current Opinion in Cell Biology 1999, 11:391-401.
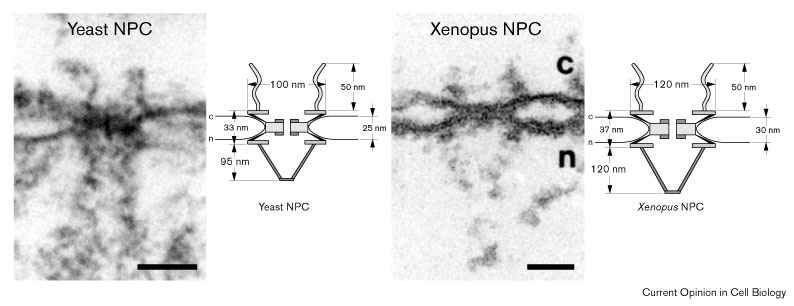
Рис. 1 Cross-sections through the NE of yeast and Xenopus oocyte nuclei (c, cytoplasm; n, nucleus), and schematic comparison of yeast and Xenopus oocyte NPCs . Scale bars 100 nm.
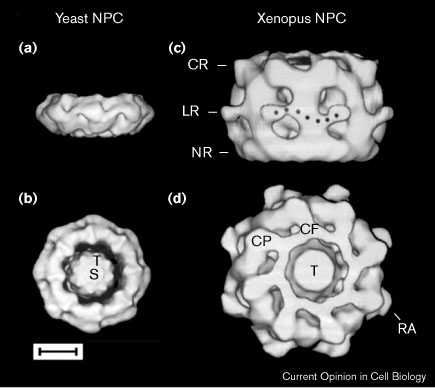
Рис. 2 A comparison between the three-dimensional structures of yeast and vertebrate NPCs. (a) Side view of the yeast NPC showing the marked difference in height relative to the Xenopus NPC and the lack of thin rings on both sides. (b) The yeast NPC as viewed from the putative cytoplasmic surface. The transporter is marked (T) and putative cargo is marked (S). (c) Side view of the Xenopus NPC reveals the cytoplasmic (CR) and nuclear (NR) thin rings that are integral parts of the spoke complex. A lumenal ring (LR) is formed by the lumenal spoke domain and the adjacent radial arms (black dots). (d) The Xenopus NPC as viewed from the cytoplasmic surface. The transporter (T) is partly obscured by a ring of collapsed cytoplasmic filaments (CF) that emanate from the cytoplasmic particles (CP). The radial arms are labeled (RA). Scale bar, 30 nm.
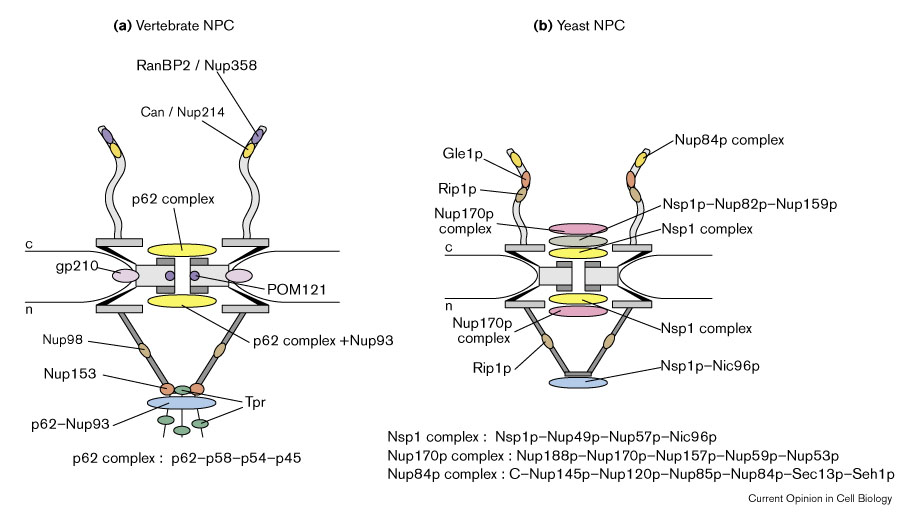
Рис. 3 Schematic diagram summarizing the immunolocalization of nucleoporin epitopes within the three-dimensional architecture of (a) the vertebrate NPC and (b) the yeast NPC. (a) In vertebrates, CAN/Nup214 and RanBP2 exhibit epitopes at the cytoplasmic fibrils [1]. The p62 complex, consisting of p62, p58, p54, and p45, exhibits epitopes at the cytoplasmic and the nuclear periphery of the central gated channel [1]; additionally, p62 exhibits an epitope at the nuclear basket. Nup93 epitopes are located at the nuclear periphery of the central gated channel and at the nuclear basket.Nup153, Nup98 and Tpr exhibit epitopes at the nuclear basket, the latter also at the intranuclear filaments. The transmembrane proteins gp210 and POM121 are predicted to have epitopes in the lumen of the NE and on the NPC proper, respectively. (b) In yeast, the nucleoporins of the Nup84p complex (i.e. C-Nup145p, Nup120p, Nup85p, Nup84p, Sec13p, and Seh1p) display epitopes at the cytoplasmic fibrils. Gle1p and Rip1p epitopes reside at the cytoplasmic fibrils, Rip1p additionally at the nuclear basket . The epitopes of the Nsp1p complex (Nsp1p–Nup49p–Nup57p–Nic96p) are located at the cytoplasmic and the nuclear periphery of the central gated channel. Moreover, Nsp1p and Nic96p show epitopes at the distal ring of the nuclear basket. Epitopes of the Nsp1p–Nup82p–Nup159p complex are displayed at the cytoplasmic periphery of the central gated channel. The Nup170p complex (i.e. Nup188p, Nup170p, Nup157p, Nup59p, and Nup53p), exhibit epitopes at the cytoplasmic and nuclear face of the NPC core, with no clear assignment to distinct substructures of the NPC. As all these localization studies have been performed with antibodies directed against epitopes of nucleoporins or tags fused to nucleoporins, however, we are only at the beginning of understanding the complete three-dimensional molecular architecture of the NPC.
The nuclear pore complex
Stephen A Adam Genome Biology 2001 2(9): reviews0007.1-0007.6
http://genomebiology.com/2001/2/9/reviews/0007
The nuclear pore complex (NPC) формируют
каналы для обмена информацией между ядром
и цитоплазмой. Структура NPC очень сходна по
виду у всех эукариот от дрожжей до
человека. Как эксклюзивные места для
переносе макромолекул между ядром и
цитоплазмой NPC являются важными
контрольными точками для регуляции
экспрессии генов.
| A comparison of the structures of (a) vertebrate
и (b) yeast nuclear pore complexes (NPCs)
Комплексы ядреных пор являются белковыми структурами,
внедренными в двойную мембрану ядерной
оболочки. Чтобы понять как функционируют NPC
необходимо изучить сходство и различия
между NPCs дрожжами Saccharomyces cerevisiae и
лягушками Xenopus laevis. NPC - это большая
структура с м.в. около 125 MDa у
позвоночных и 66 MDa у дрожжей. Ядра клеток
позвоночных содержат порядка 2,000 NPCs,
тогда как более мелкия ядра дрожжей около 200. NPCs
имеют 8-кратную ротационную симметию
относительно центральной оси поры и
двукратную зеркальную симметрию
относительно плоскости ядерной оболочки,
это указывает на то, что происходит
сборка модулярной структуры.
Помимо различий в размерах NPCs
дрожжей (yNPCs) и позвоночных (vNPCs) отличаются
по нескольким фундаментальным
структурным признакам. Основная масса vNPC
находится в виде трех-частной структуры,
которая окружает и поддерживает
центральный транспортер (Рис. 1a);
трехслойная структура состоит из тонкого
цитоплазматического и ядерного колец,
которые как в сэндвиче заключают
центральный со спицами (spoke) домен. Этот
домен также состоит из внутреннего
кольцевого элемента, который окружен
структурой, называемой central transporter и
наружным просветным кольцом, которое
взаимодействует с ядерной мембраной;
вертикальные спицевые элементы
соединяются с цитоплазматическим и
ядерным кольцами. Отходящие от
цитоплазматического тонкого кольца
выросты являются 8 филаментами, каждая из
которых 2-3 nm в диаметре и примерно 50 nm в
длину. Нуклеоплазматическая сторона NPC представлена восмью в 100 nm филаментами, которые соединены с небольшой кольцевой структурой, образуя структуру, подобную рыболовной сети, отходящей от ядерного тонкого кольца (Рис. 1a).
yNPC намного проще по структуре(Рис. 1b ). По сравнению с vNPC, они не содержат тонких цитоплазматического и ядерного кольца, и просветное кольцо центрального spoke домена. Вместо этого, центральный транспортер окружается с помощью внутреннего spoke кольца и ассоциированного с мембраной кольца. Т.к. у них отсутствует наружное тонкие кольца, то yNPCs лишены также частей spoke доменов, которые имеются в vNPCs. Подобно vNPCs, yNPCs содержит филаментозные структуры, исходящие из цитоплазматической и нуклеоплазматической сторон комплекса, хотя у дрожжей эти филаменты несколько короче, чем у позвоночных. В целом, yNPC меньше (96 nm в диаметре и 35 nm высотой), чем vNPC (145 nm диаметр и 80 nm высота), и составляет половину массы vNPC и занимает 1/5 объема.
Transport through the NPC
Показано, что NPC формирует барьер для избирательной проницаемости через ядерную оболочку. Интерные полимеры и малые белки менее 9 nm в диаметре или менее 30-40 kDa по массе м свободно диффундировать через NPC. Большие честицы проходят через NPC с помощью облегчающего механизма, который еще плохо изучен, и NPC м. приспосабливаться к транспорту частиц величиной до 25 nm в диаметре. NPCs м расширяться радиально, способствуя прхождению крупных частиц. Структура центрального транспортера образует путь для переноса макромолекул. Хотя многочисленные эксперименты показывают, что NPC формируют aqueous каналы между ядерным и цитоплазматическим компартментнами, patch-clamp техника позволила определить, что комплексы пор м. действительно быть регулируемыми каналами, которые ведут себя подобно ионным каналам, которые м. открываться и закрываться.
Основаная масса переноса макромолекул через NPC обеспечивается системой мобильных транспортных
белков, отличных от собственно NPC,
которы действуют вместе, чтобы перемещать
молекулы между ядерным и цитоплазматическим компартментом.
Мобильные транспортеры, которые
непосредственно связывают молекулы, чтобы
их транспортировать, являются членами
семейства белков, обладющими общими
свойствами, с теми, что найдены у члена
семейства importin β, известного также
как karyopherin β . Все белки мобильных
транспортеров имеют мол. массу между
90 и 130 kDa, со значительной частью каждого белка
упакованного в серию спиральных HEAT
повторовs. HEAT повторы являются тендемно
повторяющимися мотивами из 40-50
аминокислот, которые формируют интерактивные поверхности у ряда белков. Повторы HEAT транспортеров образуют два
домена, на N-терминальной половине, которые
связывают малую GTPase Ran , когда ассоциирована
с GTP и нa С-терминальной половине с доменом,
связывающим груз. Он является
доменовой структурой, которая позволяет
каждому транспортру функционировать
или как импортирующий рецептор или как
экспортирующий рецептор: экспортные
рецепторы связываются свой груз
кооперативно с Ran-GTP, тогда как
импортирующие рецепторы высвобождают
свой груз, когда связываются с Ran-GTP. Ran
является таким образом ключевым
компонентом, который определяет свойства
компартмента для транспорта. Ядерная
локализация Ran guanine-nucleotide exchange factor, RanGEF, как
полагают, содержит ядерный Ran в GTP-связанной
форме. Ran GTPase-activating protein (RanGAP)
локализуется на цитоплазматической
стороне ядерной поры, он гарантирует. что
любой Ran в цитоплазме будет связан с
GDP. Др. малые Ran-связывающие белки участвуют
в модуляции GTP гидролиза или транспорта
Ran из цитоплазмы в ядро. Увеличение
количества RanGTP-связывающих мобильных
транспортных белков в ядре подтверждается
обилием в ядре Ran .
Ключевой ступеньтю
транспорта является взаимодействие
мобильных транспортных белков с нуклеопоринами, структурными белками
комплекса пор. Эти взаимодействия
являются базой для селективного транспорта макромолекул через поры.
Большинство нуклеопоринов вовлекается
непосредственно в транспорт общих
повторов, содержащих фенилаланин-глициновые
дипептиды , и известны как FG nucleoporins. FG
повиторы взаимодействуют со специфичесими HEAT повторами мобильных
транспортеров. Однако, это довольно слабые
взаимодействия, каждый из FG нуклеопоринов
содержит множественные FG повторы, которые
должны обеспечивать сайты множественных
взаимодействий для транспортеров.
Разные транспортеры, по-видимому, следуют
разными путями через NPC, хотя некоторые из
этих путей имеют несколько общих
нуклеопоринов. У S. cerevisiae 14
транспортеров идентифицировано в геноме; 9
функционируют как импортеры, а 4 как
экспортеры. По крайней мере один
мобюильный транспортер, Kap124/Msn5, м.
выполнять двойную функцию импорта и
экспорта. но большинство специфично или к
импорту или экспорту. Имеется, по крайней
мере 22 транспортера в клетках человека,
так что некоторые из них не будут иметь
своих гомологов у дрожжей.
Как транспортеры взаимодействуют с
нуклеопоринами? В ранних моделях транслокации белков через NPC
предполагалось, что асимметричное
располжение нуклеопоринов в NPC
образует градиент сродства вдоль которого
мобильный транспортер движется за счет
диффузии, с повторяющимися ассоциациями и
диссоциациями с нуклеопоринами.
Недвание исследования сродства транспортного комплекса для
иммобилизованных нуклеопоринов подтвердили определенные аспекты этого
механизма. При более детальном протеомном анализе были идентифицированы транспортеры, а также и др. белки, которые
взаимодействуют с 8 из 13 дрожжевых FG
нуклеопоринов. Следовательно, некоторые
нуклеопорины взаимодействуют с не менее,
чем 10 различными транспортерами, тогда ка
др. нуклеопорины являюся избирательными
для Kap95p-Kap60p мобильного транпортерорго
комплекса.
Analysis of nuclear pore complex structure
и composition
Ранние подсчеты белков, составляющих коплекс поры, давали
значения между 100 и 200. Подсчеты базировались на массе пор (125 MDa), 8 радиальных складках и двух складках
зеркальной симметрии пор и подсчетах
среднего размера нуклеопоринов (100 kDa).
Рядом исследователей были идентифицированы субкомплексы
нуклеопоринов, которые м.б. локализованы в
центральном транспортере,цитплазматических филаментах и ядерных
корзинках.
Расшифровка генома дрожжей сделала
возможным идентифицировать сходные с NPC
белки по сходству последовательностей с
известными нуклеопоринами. Затем
очистка yNPC сделала возможно потенциальную
идентификацию всех белков комплекса поры
с помощью протеомного анализа. Было
установлено, что yNPC состоят примерно из 30
белков, что намного меньше первоначальных
расчетов, причем каждый белок представлен
от8 до 56 копий на пору. Локализация всех 30
белков с помощью protein-A tagging
или immunolocalization с помощью специфических
антител принесла др. сюрприз: большинство
дрожжевых нуклеопоринов довольно симметрично распределены как на
цитоплазматической, так и нуклеоплазматической стороне поры.
Только 5 нуклепоринов обнаруживались
исключительно или на ядерной или
цитоплазматической стороне комплекса
поры, тогда как другие 4 нуклеопорина имели
'biased' локализацию, предоминирующей на
одной или другой стороне. 7 из этих 9
нуклеопоринов содержат FG повторы и их
локализация на периферических структурах NPC помещает их в идеальную
позицию для взаимодействия с мобильными
транспортерами. Локализован Nsp1p
и взаимодействующие с ним нуклеопорины в
трех отдельных субкомплексах внутри NPC.
Эти результаты показали. что хотя
индивидуальные нуклеопорины распределены
по всему комплексу поры, они м.б.
представлены в разных комплексах с др.
нуклеопоринами в каждой субструктуре
комплекса поры, генерируя функциональную
асимметрию. С др. стороны, локализация
некоторых нуклеопроинов позвоночных
подтверждает, что локализация
нуклеопоринов в vNPC более симметрична, чем
в yNPC. Уже давно предполагалось. что
векторная природа импорта и экспорта
обусловлна частично асимметричным
распределением нуклеопроинов.
Является ли эта асимметрия необходимой
только для захвата (docking) транспортных
комплексов в порах или также необходима
для транслокации через пору, остается
предметом споров.
Evolutionary conservation of nuclear pore complexes
Помимо структурных различий (Рис.1)
существуют и функциональные различия yNPCs
и vNPCs. Ядерная оболочка у дрожже не имет
ядерной ламины, промежуточной филаменты
содержащей структуры, лежащей в основе
ядерной мембраны; имеются данные.
указывающие на сущестование взаимодействий между NPC
и ламиной в др. клетках. Клетки почкующихся
дрожжей не нарушают ядерной оболочки при
митозах и не ведут к разборке NPCs как это
происходит у позвоночных. Эти два
структурных различия скорее всего
отражаются на белковом составе пор у
клеток двух типов. Структурный анализ vNPCs
и yNPCs указывает на то, что они взаимодействуют по-разному с ядерной
мембраной. NPCs закреплены в ядерной
мембране с помощью интегральных мембранных белков. 4 таkих белка
присутствуют в yNPC, тогда как только два
идентифицированы у vNPC; и причем ни один из двух белков
позвoноночных не родственен белкам
дрожжей. Различия в способе закрепления
NPCs в мембранах скорее всего связаны с
динамикой NPC во время клеточного цикла.
Идентификация нуклеопоринов позвоночных осложнена. Тесная
ассоциация vNPC с подлежащей ламиной
ведет к тому, что множество белков не
относящихся к ядерной оболочке
присутствуют в любом препарате. Недавно
удалось преодолеть эту проблему,
используя специализированные структуры .
называемые annulate lamellae, которые
присутствуют в клетках некоторых метазоа.
Annulate lamellae это уплощенные мешки из двойных
мембран с плотно упакованными радами
комплексов пор, которые обнаруживаются в
дифференцирующихся и быстро растущих
клетках. Эти структуры м. собирать in vitro
в больших количествах из экстрактов яиц Xenopus
и м. очищать. Оказалось. что большинство vNPCs
содержит 40-50 индивидуальных видов белков,
количество сходное с таковым у yNPC несмотря
на большие размеры vNPC.
Когда известные дрожжевые нуклеопорины
сравнили с позвоночными, то оказалось
удивительно мало гомологии между белками.
Только около половины нуклеопоринов
позвночных имели явную гомологию с
дрожжевыми по последовательностя, функции
и локализации. Некоторые имели более
слабое и очень слабоe сходство
последовательностей. В большинстве
случаев, единственным сходством между
белками было присутствие FG повторов. По
крайней мере два нуклеопорина, Nup153
и Nup358, содержат домены цинковые пальчики,
тогда как ни один из них не найден у
нуклеопоринов дрожжей. Др. существенным
отличием было отсутствие дрожевых
нуклеопоринов, содержащих Ran-связывающие
домены. Nup2p единственный дррожжевой
нуклеопорин с Ran-связывающим доменом,
однако его роль как аутентичного
нуклеопорина еще под вопросом. vNPCs, с др.
стороны, содержат по крайней мере два Ran-связывающих
нуклеопорина, Nup358 и Nup153, оба участвуют в инициальных и
терминальных событиях импорта и экспорта.
Являются ли эти различия фундаментальными
для механизмов транспорта, неизвестно.
Теперь, когда все дрожжевые нуклеопорины,
кодируемые в геноме, идентифицированы, то
взаимодействия между ними внутри NPC м.б.
картированы биохимичесикими методами.
NPC субкомплесы были идентифицированы в
vNPCs, однако внесение меченных (tagged) или
модифицированных нукелеопротеинов в NPC
ограничено повторной сборкой из яйцевых
экстрактов Xenopus. Недавно была
предолена и эта трудность, предложен
двухступенчатый 'organelle trap' подход.
Растворимые белки из яйцевых экстрактов Xenopus
пропускали через affinity column , содержащий
интересующий лиоганд. Связанные белки
элюировали метили биотином, и воссоздавали в NPCs annulate lamellae, собираемых in vitro. Используя этот
подход идентифицирован новый нуклеопорин
позвоночных, Nup188, он комплексуется с двумя
другими уже изсестными нуклеопоринами;
оба имеют дрожжевых гомологов и оба
дрожжевые гомологи образуют комплекс с
дрожжевым Nup188. Хотя yNup188 и vNup188 имеют ограниченное сходство
последовательностей, но их присутствие в
сходных комплексах указывает на то. что
они и ассоциируемые с ними белки формируют
законсервированный субкомплекс внутри NPC.
Структура NPC у Chironomus обнаружила
занчительную консервацию структур у
беспозвоночных и позвоночных. Простое
изучение Flybase и WormPD баз данных выявило.
что только около десяти гомологос
нуклеопоринов идентифицировано у каждого
из этих организмов. Недавно у C. elegans с
помощзью моноклональных антителраспознан Nup62p
млекопитающих и несколько др. нуклеопринов. По крайней мере Nup62p м.б. законсервирован. Геномика открывает новые
возможности для сравнения геномов Drosophila и C. elegans для анализа NPC.