Члены семейства сигнальных молекул Wnt играют разные важные роли в развитии и болезнях. Идентифицировано Frizzled семейство белков с 7 трансмембранными доменами в качестве рецепторов Wnt[1][2][3][4][5] (Fig. 1).
Bhanot P, Brink M, Samos CH, Hsieh JC, Wang Y, Macke JP, Andrew D, Nathans J, Nusse R: A new member of the frizzled family from Drosophila functions as a Wingless receptor. Nature 1996, 382: 225–230.
Wang YS, Macke JP, Abella BS, Andreasson K, Worley P, Gilbert DJ, Copeland NG, Jenkins NA, Nathans J: A large family of putative transmembrane receptors homologous to the product of the Drosophila tissue polarity gene frizzled. J Biol Chem 1996, 271: 4468–4476.
Cadigan KM, Nusse R: Wnt signaling: a common theme in animal development. Genes Dev 1997, 11: 3286–3305.
Moon RT, Brown JD, Torres M: WNTs modulate cell fate and behaviour during vertebrate development. Trends Genet 1997, 13: 157–162.
Wodarz A, Nusse R: Mechanisms of Wnt signaling in development. Annu Rev Cell Dev Biol 1998, 14: 59–88.
Giarre M, Semenov MV, Brown AMC: Wnt signaling stabilizes the dual-function protein beta-catenin in diverse cell types. Ann NY Acad Sci 1998, 857: 43–55.
Eastman Q, Grosschedl R: Regulation of LEF-1/TCF transcription factors by wnt and other signals. Curr Opin Cell Biol 1999, 11: 233–240.
• Fagotto F, Jho E, Zeng L, Kurth T, Joos T, Kaufmann C, Costantini F: Domains of axin involved in protein–protein interactions, Wnt pathway inhibition, and intracellular localization. J Cell Biol 1999, 145: 741–756.
Kitagawa M, Hatakeyama S, Shirane M, Matsumoto M, Ishida N, Hattori K, Nakamichi I, Kikuchi A, Nakayama K, Nakayama K: An F-box protein, FWD1, mediates ubiquitin-dependent proteolysis of beta-catenin. EMBO J 1999, 18: 2401–2410.
Hart M, Concordet JP, Lassot I, Albert I, del los Santos R, Durand H, Perret C, Rubinfeld B, Margottin F, Benarous R, Polakis P: The F-box protein beta-TrCP associates with phosphorylated beta-catenin and regulates its activity in the cell. Curr Biol 1999, 9: 207–210.
Liu C, Kato Y, Zhang Z, Do VM, Yankner BA, He X: beta-Trcp couples beta-catenin phosphorylation-degradation and regulates Xenopus axis formation. Proc Natl Acad Sci USA 1999, 96: 6273–6278.
Smalley MJ, Sara E, Paterson H, Naylor S, Cook D, Jayatilake H, Fryer LG, Hutchinson L, Fry MJ, Dale TC: Interaction of Axin and Dvl-2 proteins regulates Dvl-2-stimulated TCF-dependent transcription. EMBO J 1999, 18: 2823–2835.
Kaletta T, Schnabel H, Schnabel R: Binary specification of the embryonic lineage in Caenorhabditis elegans. Nature 1997, 390: 294–298.
•• Lin R, Hill RJ, Priess JR: POP-1 and anterior-posterior fate decisions in C. elegans embryos. Cell 1998, 92: 229–239.
Thorpe CJ, Schleisinger A, Carter JC, Bowerman B: Wnt signalling polarizes an early C. elegans blastomere to distinguish endoderm from mesoderm. Cell 1997, 90: 695–705.
Rocheleau CE, Downs WD, Lin R, Wittmann C, Bei Y, Cha YH, Ali M, Priess JR, Mello CC: Wnt signaling and an APC-related gene specify endoderm in early C. elegans embryos. Cell 1997, 90: 707–716.
Bullions LC, Levine AJ: The role of beta-catenin in cell adhesion, signal transduction, and cancer. Curr Opin Oncol 1998, 1: 81–87.
Nishimura M, Yu G, Levesque G, Zhang DM, Ruel L, Chen F, Milman P, Holmes E, Liang Y, Kawarai T et al.: Presenilin mutations associated with Alzheimer disease cause defective intracellular trafficking of beta-catenin, a component of the presenilin protein complex. Nat Med 1999, 5: 164–169.
Levesque G, Yu G, Nishimura M, Zhang DM, Levesque L, Yu H, Xu D, Liang Y, Rogaeva E, Ikeda M, Duthie M et al.: Presenilins interact with armadillo proteins including neural-specific plakophilin-related protein and beta-catenin. J Neurochem 1999, 72: 999–1008.
Tao YS, Edwards RA, Tubb B, Wang S, Bryan J, McCrea PD: beta-Catenin associates with the actin-bundling protein fascin in a noncadherin complex. J Cell Biol 1996, 134: 1271–1281.
Gallet A, Erkner A, Charroux B, Fasano L, Kerridge S: Trunk-specific modulation of wingless signalling in Drosophila by teashirt binding to armadillo. Curr Biol 1998, 8: 893–902.
• Gallet A, Angelats C, Erkner A, Charroux B, Fasano L, Kerridge S: The C-terminal domain of Armadillo binds to hypophosphorylated Teashirt to modulate Wingless signalling in Drosophila. EMBO J 1999, 18: 2208–2217.
Torres MA, Eldar-Finkelman H, Krebs EG, Moon RT: Regulation of ribosomal S6 protein kinase-p90(rsk), glycogen synthase kinase 3, and beta-catenin in early Xenopus development. Mol Cell Biol 1999, 19: 1427–1437.
Cohen P: The Croonian Lecture 1998. Identification of a protein kinase cascade of major importance in insulin signal transduction. Philos Trans R Soc Lond B Biol Sci 1999, 354: 485–495.
Kim E, Arnould T, Sellin LK, Benzing T, Fan MJ, Gruning W, Sokol SY, Drummond I, Walz G: The polycystic kidney disease 1 gene product modulates Wnt signaling. J Biol Chem 1999, 274: 4947–4953.
Larabell CA, Torres M, Rowning BA, Yost C, Miller JR, Wu M, Kimelman D, Moon RT: Establishment of the dorso-ventral axis in Xenopus embryos is presaged by early asymmetries in beta-catenin that are modulated by the Wnt signaling pathway. J Cell Biol 1997, 136: 1123–1136.
Medina A, Wendler SR, Steinbeisser H: Cortical rotation is required for the correct spatial expression of nr3, sia and gsc in Xenopus embryos. Int J Dev Biol 1997, 41: 741–745.
•• Novak A, Hsu SC, Leung-Hagesteijn C, Radeva G, Papkoff J, Montesano R, Roskelley C, Grosschedl R, Dedhar S: Cell adhesion and the integrin-linked kinase regulate the LEF-1 and beta-catenin signaling pathways. Proc Natl Acad Sci USA 1998, 95: 4374–4379.
Papkoff J, Aikawa M: WNT-1 and HGF regulate GSK3 β activity and β-catenin signaling in mammary epithelial cells. Biochem Biophys Res Commun 1998, 247: 851–858.
Kengaku M, Capdevila J, Rodriguez-Esteban C, De La Pena J, Johnson RL, Belmonte JCI, Tabin CJ: Distinct WNT pathways regulating AER formation and dorsoventral polarity in the chick limb bud. Science 1998, 280: 1274–1277.
Staal FJ, Burgering BM, van de Wetering M, Clevers HC: Tcf-1-mediated transcription in T lymphocytes: differential role for glycogen synthase kinase-3 in fibroblasts and T cells. Int Immunol 1999, 11: 317–323.
• Slusarski DC, Corces VG, Moon RT: Interaction of Wnt and a Frizzled homologue triggers G-protein-linked phosphatidylinositol signalling. Nature 1997, 390: 410–413
•• Meneghini MD, Isnitani T, Clayton Carter J, Hisamoto N, Ninomiya-Tsuji J, Thorpe CJ, Hamill DR, Matsumoto K, Bowerman B: MAP kinase and Wnt pathways converge to down-regulate an HMG domain repressor in C. elegans. Nature 1999, 399: 793–798.
•• Rochleau C, Yasuda J, Shin TH, Lin R, Sawa H, Okano H, Priess JR, Davis RJ, Mello CC: WRM-1 activates the LIT-1 protein kinase to transduce anterior/posterior polarity signals in C. elegans. Cell 1999, 97: 717–726.
•• Wesley C: Notch and Wingless regulate expression of cuticle patterning genes. Mol Cell Biol 1999, in press.
• Piccolo S, Agius E, Leyns L, Bhattacharyya S, Grunz H, Bouwmeester T, De Robertis EM: The head inducer Cerberus is a multifunctional antagonist of Nodal, BMP and Wnt signals. Nature 1999, 397: 707–710.
• Hsieh JC, Kodjabachian L, Rebbert ML, Rattner A, Smallwood PM, Samos CH, Nusse R, Dawid IB, Nathans J: A new secreted protein that binds to Wnt proteins and inhibits their activities. Nature 1999, 398: 431–436.
Tsuda M, Kamimura K, Nakato H, Archer M, Staatz W, Fox B, Humphrey M, Olson S, Futch T, Kaluza V et al.: A cell surface proteoglycan, Dally, regulates Wingless signalling in Drosophila. Nature 1999, in press.
• Klein T, Martinez Arias A: The Vestigial gene product provides a molecular context for the interpretation of signals during the development of the wing in Drosophila. Development 1999, 126: 913–925.
Bejsovec A, Martinez Arias A: Roles of Wingless in patterning the larval epidermis of Drosophila. Development 1991, 113: 471–485.
Heemskerk J, DiNardo S, Kostriken R, O'Farrell PH: Multiple modes of engrailed regulation in the progression towards cell fate determination. Nature 1991, 352: 404–410.
Garcia-Garcia MJ, Ramain P, Simpson P, Modolell J: Different contributions of pannier and wingless to the patterning of the dorsal mesothorax of Drosophila. Development 1999, in press.
Sampedro J, Johnston P, Lawrence PA: A role for wingless in the segmental gradient of Drosophila? Development 1993, 117: 677–687.
Martinez Arias A: Interactions between Wingless and Notch during the assignation of cell fates in Drosophila. Int J Dev Biol 1998, 42: 325–333.
Baylies MK, Martinez Arias A, Bate M: wingless is required for the formation of a subset of muscle founder cells during Drosophila embryogenesis. Development 1995, 121: 3829–3837.
Cui Y, Tian Q, Christian JL: Synergistic effects of Vg1 and Wnt signals in the specification of dorsal mesoderm and endoderm. Dev Biol 1996, 180: 22–34.
Hoppler S, Moon RT: BMP-2/-4 and Wnt-8 cooperatively pattern the Xenopus mesoderm. Mech Dev 1998, 71: 119–129.
Giese K, Kingsley C, Kirshner J, Grosschedl R: Assembly of a TCRalpha enhancer complex is dependent on LEF-1 induced DNA bending and multiple protein-protein interactions. Genes Dev 1995, 9: 995–1008.
•• Hsieh JC, Rattner A, Smallwood PM, Nathans J: Biochemical characterization of Wnt-frizzled interactions using a soluble, biologically active vertebrate Wnt protein. Proc Natl Acad Sci USA 1999, 96: 3546–3551.
• Kennerdell JR, Carthew RW: Use of dsRNA-mediated genetic interference to demonstrate that frizzled and frizzled 2 act in the wingless pathway. Cell 1998, 95: 1017–1026.
• Muller H, Samanta R, Wieschaus E: Wingless signaling in the Drosophila embryo: zygotic requirements and the role of the frizzled genes. Development 1999, 126: 577–586.
• Bhat K: frizzled and frizzled 2 play a partially redundant role in wingless signaling and have similar requirements to wingles in neurogenesis. Cell 1998, 95: 1027–1036.
Xu Q, D'Amore PA, Sokol SY: Functional and biochemical interactions of Wnts with FrzA, a secreted Wnt antagonist. Development 1998, 125: 4767–4776.
Nasevicius A, Hyatt T, Kim H, Guttman J, Walsh E, Sumanas S, Wang Y, Ekker SC: Evidence for a frizzled-mediated wnt pathway required for zebrafish dorsal mesoderm formation. Development 1998, 125: 4283–4292.
Deardorff MA, Tan C, Conrad LJ, Klein PS: Frizzled-8 is expressed in the Spemann organizer and plays a role in early morphogenesis. Development 1998, 125: 2687–2700.
Leyns L, Bouwmeester T, Kim SH, Piccolo S, DeRobertis EM: Frzb-1 is a secreted antagonist of Wnt signaling expressed in the Spemann organizer. Cell 1997, 88: 747–756.
Wang SW, Krinks M, Lin KM, Luyten FP, Moos M: Frzb, a secreted protein expressed in the Spemann organizer, binds and inhibits Wnt-8. Cell 1997, 88: 757–766.
Saldanha J, Singh J, Mahadevan D: Identification of a Frizzled-like cysteine rich domain in the extracellular region of developmental receptor tyrosine kinases. Protein Sci 1998, 7: 1632–1635.
Dierick HA, Bejsovec A: Functional analysis of Wingless reveals a link between intercellular ligand transport and dorsal-cell-specific signaling. Development 1998, 125: 4729–4738.
Bejsovec A, Wieschaus E: Signaling activities of the Drosophila wingless gene are separately mutable and appear to be transduced at the cell surface. Genetics 1995, 139: 309–320.
Reichsman F, Smith L, Cumberledge S: Glycosaminoglycans can modulate extracellular localization of the Wingless protein and promote signal transduction. J Cell Biol 1996, 135: 819–827.
Haerry TE, Heslip TR, Marsh JL, O'Connor MB: Defects in glucuronate biosynthesis disrupt Wingless signaling in Drosophila. Development 1997, 124: 3055–3064.
Binari RC, Staveley BE, Johnson WA, Godavarti R, Sasisekharan R, Manoukian AS: Genetic evidence that heparin-like glycosaminoglycans are involved in wingless signaling. Development 1997, 124: 2623–2632.
Hacker U, Lin X, Perrimon N: The Drosophila sugarless gene modulates Wingless signaling and encodes an enzyme involved in polysaccharide biosynthesis. Development 1997, 124: 3565–3573.
Glinka A, Wu W, Delius H, Monaghan AP, Blumenstock C, Niehrs C: Dickkopf-1 is a member of a new family of secreted proteins and functions in head induction. Nature 1998, 391: 357–363.
Aravind L, Koonin EV: A colipase fold in the carboxy-terminal domain of the Wnt antagonists-the Dickkopfs. Curr Biol 1998, 8: R477–R478.
Reichsman F, Moore HM, Cumberledge S: The C-terminal domain of Wingless shares significant sequence homology with a lipid-binding domain in secreted Phospholipase A2. Curr Biol 1999, 9: R353.
Artavanis-Tsakonas S, Rand MD, Lake RJ: Notch signaling: cell fate control and signal integration in development. Science 1999, 284: 770–776
Couso JP, Martinez Arias A: Notch is required for wingless signaling in the epidermis of Drosophila. Cell 1994, 79: 259–272.
Brennan K, Tateson R, Lewis K, Martinez Arias A: A functional analysis of Notch mutations in Drosophila. Genetics 1997, 147: 177–188.
Axelrod JD, Matsuno K, Artavanis Tsakonas S, Perrimon N: Interaction between Wingless and Notch signaling pathways mediated by Dishevelled. Science 1996, 271: 1826–1832.
Hartenstein V, Younossi-Hartenstein A, Lekven A: Delamination and division in the Drosophila neurectoderm: spatiotemporal pattern, cytoskeletal dynamics, and common control by neurogenic and segment polarity genes. Dev Biol 1994, 165: 480–499.
Carmena A, Gisselbrecht S, Harrison J, Jimenez F, Michelson AM: Combinatorial signaling codes for the progressive determination of cell fates in the Drosophila embryonic mesoderm. Genes Dev 1998, 12: 3910–3922.
•• Brennan K, Baylies M, Martinez Arias A: Repression by Notch is required before Wingless signalling during muscle progenitor cell development in Drosophila. Curr Biol 1999, 9: 707–710.
• Boutros M, Paricio N, Strutt DI, Mlodzik M: Dishevelled activates JNK and discriminates between JNK pathways in planar polarity and wingless signaling. Cell 1998, 94: 109–118.
• Li L, Yuan H, Xie W, Mao J, Caruso AM, McMahon A, Sussman DJ, Wu D: Dishevelled proteins lead to two signaling pathways. Regulation of LEF-1 and c-Jun N-terminal kinase in mammalian cells. J Biol Chem 1999, 274: 129–134.
Yamaguchi K, Shirakabe K, Shibuya H, Irie K, Oishi I, Ueno N, Taniguchi T, Nishida E, Matsumoto K: Identification of a member of the MAPKKK family as a potential mediator of TGF-beta signal transduction. Science 1995, 270: 2008–2011.
Choi KW, Benzer S: Rotation of photoreceptor clusters in the developing Drosophila eye requires the nemo gene. Cell 1994, 78: 125–136.
Cavallo RA, Cox RT, Moline MM, Roose J, Polevoy GA, Clevers H, Peifer M, Bejsovec A: Drosophila Tcf and Groucho interact to repress Wingless signalling activity. Nature 1998, 395: 604–608.
Roose J, Molenaar M, Peterson J, Hurenkamp J, Brantjes H, Moerer P, van de Wetering M, Destree O, Clevers H: The Xenopus Wnt effector XTcf-3 interacts with Groucho-related transcriptional repressors. Nature 1998, 395: 608–612.
• Zecchini V, Brennan K, Martinez-Arias A: An activity of Notch regulates JNK signalling and affects dorsal closure in Drosophila. Curr Biol 1999, 9: 460–469
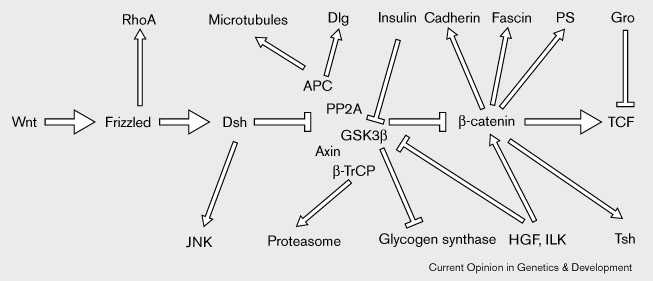
Figure 1 Основной канонический путь передачи сигналов Wnt (представлен горизонтальными стрелками)и его связь с другими процессами и путями. Wnts сыязываются Frizzled рецепторами и каким-то образом передают сигнал Dishevelled (Dsh). Кроме того некоторые Drosophila Frizzled белки взаимодействуют с RhoA во время возникновения полярности ткани, тогда как Dishevelled (Dsh) может регулировать активность Jun amino-terminal kinase (JNK) [75••][76•]. Центральным свойством передачи сигналов Wnt является модуляция стабильности и локализация β-catenin посредством комплекса, котороый включает Axin, APC, PP2A, and GSK3β. Имеется, по крайней мере, 3 пула β-catenin: цитозольный, ядерный, и связанный с мембраной через взаимодействие с Cadherins. β-catenin может также взаимодействовать с Presenilins (PS), actin-bundling белком Fascin, и Drosophila Zn finger protein Teashirt (Tsh). Когда цитозольный β-catenin становится фосфорилированным в базирующемся на Axin комплексе, то он связывается с β-TrCP, который затем осуществляет его протеолиз. APC, важный компоенент Axin-based комплекса, он взаимодействует с микротрубочками и и с Drosophila белком супрессирующим опухоли Discs large (Dlg). В ответ на сигналы Wnt, цитозольный β-catenin стабилизируется, если Dsh инактивирует GSK3β в Axin комплексе. Повышение концентрации цитозольного β-catenin сопровождается транслокацией β-catenin в ядро, где он формирует комплекс с членами семейства TCF транскрипционных факторов. В дополнение к сигналам Wnt, hepatocyte growth factor (HGF) и integrin-linked kinase (ILK) могут ингиибровать GSK3β и способстовать образованию комплекса β-catenin/TCF [28••][29]. GSK3β участвует также в метаболизме гликогена и регулируется инсулином. Помимо связывания β-catenin, TCFs может ассоциировать с транскрипционным ко-репрессором Groucho (Gro). Важной функцией передачи сигналов Wnt м.б. изменение функции TCF из транскрипционного репрессора, когда он связан с Gro, в активатор, когда он соединяется с β-catenin> Figure 1 Flow chart of the core of the canonical Wnt signalling pathway (represented by horizontal arrows) and its links with other processes and pathways through interactions of individual components. Wnts bind to Frizzled receptors and somehow transmit a signal to Dishevelled (Dsh). In addition to their roles in Wnt signalling, some Drosophila Frizzled proteins interact with RhoA during the establishment of tissue polarity, while Dishevelled (Dsh) can regulate the activity of the Jun amino-terminal kinase (JNK) [75••][76•]. A central feature of Wnt signaling is modulation of the stability and localization of β-catenin via a complex that includes Axin, APC, PP2A, and GSK3β. There are at least three pools of β-catenin: cytosolic, nuclear, and membrane-bound via interactions with Cadherins. β-catenin can also interact with Presenilins (PS), the actin-bundling protein Fascin, and the Drosophila Zn finger protein Teashirt (Tsh). When cytosolic β-catenin becomes phosphorylated in the Axin-based complex, it is bound by β-TrCP which then triggers its proteolysis. APC, an important component of the Axin-based complex, has also been shown to interact with microtubules and with the Drosophila tumor suppressor protein Discs large (Dlg). In response to Wnt signalling, cytosolic β-catenin is stabilized when Dsh inactivates GSK3β in the Axin complex. Increased abundance of cytosolic β-catenin is followed by translocation of β-catenin to the nucleus where it forms complexes with members of the TCF family of transcription factors. In addition to Wnt signalling, hepatocyte growth factor (HGF) and integrin-linked kinase (ILK) can inhibit GSK3β and promote β-catenin/TCF complex formation [28••][29]. GSK3β also functions in glycogen metabolism and is regulated by insulin. Apart from binding β-catenin, TCFs can associate with the transcriptional co-repressor Groucho (Gro). An important function of Wnt signalling may be to change the function of TCF from a transcriptional repressor, when it is bound to Gro, to an activator when it is bound to β-catenin.
Центральным свойством этого пути является Wnt-обусловленная модуляция стабильности цтозольного пула β-catenin [6] и взаимодействие β-catenin с членами lymphoid enhancer factor 1 (LEF1)/T-cell factor (TCF) семейства транскрипционных факторов (reviewed in [7]). В отсутствие Wnt сигналов, цитозольный пул β-catenin подвергается активной деградации после того как он становится фосфорилированным благодаря ассоциации с большим комплексом, в который включен Axin, adenomatous polyposis coli (APC) белок, protein phosphatase 2A (PP2A), glycogen synthase kinase 3β (GSK3β) и β-transducin repeat containing protein (β-TrCP) [8•][9][10][11]. Детальный анализ этого процесса указывает на то, что Dishevelled также часть комплекса [8•][12]. Связываение Wnt белков рецепторами Frizzled активирует Dishevelled, который ингибирует киназу GSK3β в Axin-based комплексе, и делает возможным накопление в цитоплазме стабилизированного β-catenin. β-catenin затем поступает в ядро и связывается с с членами семейства LEF1/TCF. у Caenorhabditis elegans, этот процесс ведет к подавлению транскрипционной репрессии, обусловливаемой с помощью члена семейства TCF - POP-1 [13][14••][15][16]. У Drosophila и позвоночных, однако, β-catenin ,по-видимому, конвертирует TCF из транскрипционного репрессора в активатор (reviewed in [7]).
В действительности путь передачи сигнала усложняется функциональными ответвлениями, так как большинство его компонентов не только участвует в передаче Wnt сигнала, но и участвует в других биохимических или клеточных процессах(Fig. 1). Напр., β-catenin играет важную роль в клеточной адгезии [17] и взаимодействует также с Presenilins [18][19], actin-bundling белком Fascin [20] и с Drosophila гомеозисным кофактором Teashirt [21][22•]. Другой компонент пути, GSK3β, участвует в передаче сигналов инсулина и в метаболизме гликогена [23][24]. Имеются условия. при которых нижестоящие эффекты сигнального Wnt пути могут запускаться без Wnts [25][26][27][28••][29] and others in which certain Wnts can signal without apparently involving some of the core components of the canonical signalling pathway [30][31][32•].
Эти наблюдения — вместе с рядом находок
о молекулах, вовлекаемых в передачу
сигналов Wnt [33••][34••][35••],
и о белках, с которыми Wnts могут
взаимодействовать во внеклеточных
пространствах [36••][37•][38•][39] —
подтверждает необходимость
обновления точки зрения на передачу
сигналов Wnt .
Функции Wnts
в развитии
Исследования с
потерей функции генов Wnt выявили
высокую степень плейотропии [3][4].
Кажется, что Wnt участвует во многих, ели не
большинстве процессов развития.
Обратимся
к wingless, первому выявленному
у Drosophila Wnt гену. Анализ его роли
в развитии и формировании паттерна
крыльев [40•]
показал, что он является фактором,
обеспечивающим гарантии, что клетка
получит инструкции, предоставляемые
другими сигналами скорее, чем сам
предписывает такие инструкции.
Пермиссиваня функция Wingless
сравнима таже с его ролью в других
процессах, таких как регуляция генов engrailed
и achaete. Wingless действует не путем
инициации их транскрипции, но
стабилизируя их экспрессию внутри
доменов транскрипционой
компетентности, определяемых
соответствующими индукторами [41][42][43].
Однако Wingless может быть и
инструктивным в некоторых
примерах (for review, see [3]),
пермиссивная функция проявляется в
эктодерме и мезодерме [44][45][46].
Пермиссиваня
функция передачи сигналов Wnt
обнаруживается во время раннего
развития Xenopus и C. elegans . У Xenopus,
передача сигналов Wnt играет важную
роль в возникновении и формировании
паттерна мезодермы . В частности,
передача сигналов, активированная
эксприментально с помощью XWnt-8 ,
кооперирует с Vg1 в спецификации
мезодермы и энтодермы [47],
и с BMP-2/4 в дальнейшем паттернировании
ткани [48]. Wnt
сигналы гарантируют, что сигналы от Vg1
и BMP-2/4 стабильно регистрируются
клетками-мишенями. У C. elegans,
нижестоящие эффекторы сигнального
пути Wnt необходимы клеткам для
правильного восприятия
предназначения во время
нескольких бинарных решений, которые
сопровождают деления клеток вдоль
передне-задней оси [13][14••].
Wnt сигналы действуют как on/off
переключатели для адаптации клетками
судеб, обеспечиваемых другими
программами.
Исследования ядерных
эффекторов передачи сигналов Wnt
на молекулярном уровне позволяют
пролить свет на механизмы, лежащие в
основе процесса стабилизации иных
программ. Во время развития T-клеток
транскрипционные факторы LEF1/TCF не
активируют транскрипцию независимо, а
работают путем создания
специфических конфигураций хроматина,
которые разрешают активность других
ДНК-связывающих белков. [49].
Это свойство белков LEF1/TCF может
быть общераспространенным и
сравнимым с пермиссивной функцией Wnt
сигналов в предопределнии судьбы
клеток. В ходе развития специфические
промоторы нуждаются в правильном
расположении множественных факторов,
необходимых для их активности. В
случае Wnt-regulated промоторов
стабильность и функция этого ансамбля
белков может нуждаться в действии
LEF1/TCF , обеспечиваемого сигналами Wnt. Внеклеточные
молекулы, интегрирующие с Wnt
Белки
Wnt взаимодействуют с несколькими
различными внеклеточными или cell-surface
молекулами. По отношению к лиганд-рецептор
взаимодействиям имеются четкие
доказательства, что
внеклеточный cysteine rich domain (CRD)
белка Frizzled может связывать Wnts [50••];
исследования культур клеток [1],
генетический анализ мух [51•][52•][53•]
и эксперименты по экспрессии у
лягушек и рыб [54][55][56]
указывают на то, что это
функциональное взаимодействие.
Идентифицировано семейство
секретируемых Frizzled-related proteins (sFRPs),
члены которого действуют как
конкурентные игибиторы передачи
сигналов Wnt [57][58].
Однако , CRD мотив присутствует также в
некоторых внеклеточных белках,
которые неродственны Frizzled [59],
а Wnts взаимодействуют с другими
внеклеточнми молекулами или
рецепторами, которые не содержат CRD
доменов (Fig. 2).
Это вместе с анализом некоторых
мутантных Wingless белков ([60][61]; H Moore, S Cumberledge, A Martinez Arias, AMC Brown unpublished observations),
открывает возможность, что
внутриклеточная передача сигналов,
обеспечиваемая Wnt белками может
иметь более одного входа (input), и вообще
нуждаться в ансамбле белков на
клеточной поверхности.
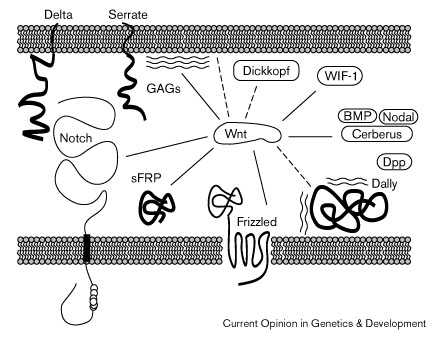
Figure 2
Сеть молекулярных взаимодействий Wnt белков во
внеклеточном пространстве.
Сплошные линии указывают на
взаимодействия, которые
задокументированы на молекулярном
уровне. Пунктирные линии указывают
на взаимодействия, выявляемые в
функциональных и,или генетических
исследованиях. Wnt белки могут
взаимодействовать с внеклеточными
доменами двух рецепторов , Frizzled и Notch. Notch
взаимодействет также с Delta и Serrate.
Связывание Wnts с Frizzled белками
происходит посредством Frizzled CRD
домена. Wnt белки могут также
взаимодействовать с sFRP/FrzBs,
секретируемыми белками,
содержащими CRD-related домен. Dickkopf, WIF-1
и Cerberus являются дальнейшими
секретируемыми белками, которые
могут взаимодействовать с Wnts. Cerberus
взаимодействует также с BMP и Nodal.
Кроме того, Wnt белки могут
взаимодействовать с glycosaminoglycans (GAGs),
которые м.б. компонентами
протеогликанов, таких как Dally. Dally
взаимодействует также с членами
семейства Drosophila BMP Decapentaplegic (Dpp).
Сходство последовательностей С-конца
Wingless и законсервированного домена
фосфолипаз указывает на то, что
Wnt белки могут также взаимодейтвовать с липидами.
Растут доказательства того, что
гликозамингликаны необходимы для
передачи сигналов Wnt [62][63][64][65]
и это, по-видимому, объясняет
наблюдение, что Drosophila
протеоглаикан Dally является
регулятором передачи сигналов Wingless
([39]; X Lin, N Perrimon personal communication). Wnts
взаимодействуют также с
секретируемыми белками, у которых
отсутствует домен CRD , таким
как Wnt inhibitory factor-1 (WIF-1) [38•], Dickkopf [66]
и Cerberus [37•] (Fig. 2). WIF-1
новый белок, содержащий повторы epidermal growth factor,
которые связываются с Wnts и
ингибируют их активность [38•]. Dickkopf
, который также взаимодействует с
передачей сигналов Wnt у Xenopus[66],
имеет С-терминальный домен, который
гомологичен colipases [67].
Это указывает на вовлечение липидов в
передачу сигналов Wnt , С-конец Wingless
обнаруживает гомологию с липид
связывающим доменом , присутсвующим у
некоторых фосфолипаз [68].
Следовательно. возможно, что Dickkopf
может нарушать передачу сигналов Wnt
модулируя его ассоциацию с липидами.
Cerberus еще один антогонист сигнализации Wnt
он м. действовать, секвестрируя Wnts
во внеклеточном пространстве.
Интересно, что Cerberus м. также
связываться и выступать антогонистом
по отношению bone morphogenetic proteins (BMPs) и Nodal [37•].
Так как сайты связывания на Cerberus
для Wnts и BMP перекрываются, то возможно.
что эти лиганды будут конкурировать
за Cerberus. Следовательнго, локальная
концентрация Cerberus и относительные
концентрации лигандов могут вносить
свой вклад во внеклеточный баланс
сигнальных активностей Wnt и BMP.
Другим белком, с которым Wingless ассоциирует
на клеточной поверхности, является Notch [36••] (Fig. 2). Notch
это большой с одиночным
трансмембранным доменом рецептор,
который играет фундаментальную роль в
латеральной ингибиции сигнализации
во время предопределения судьбы
клеток. Этот сигнал от Notch
запускается с помощью лигандов Delta/Serrate/LAG-2
семейства и обеспечивается
отщеплением внутриклеточного домена Notch
и его ассоциацией с CBF/Su(H) (C-promoter binding factor/Suppressor of Hairless)
белком (reviewed in [69]).
Связывание Wingless с Notch может в
принципе модулировать эту
активность [36••][70][71].
Дальнейшее подтверждение
взаимодействия сигнальных механизмов Wingless
и Notch получены в результате наблюдения,
что Dishevelled может связывать
внутриклеточный домен Notch и
влиять на некоторые его сигнальные
способности [72].
Потеря сигнализации Wingless
обусловливает неспособность
некоторых нейральных и мышечных
предшественников развиваться [45][46][73][74].
Этот фенотип контрастирует с
избыточным количеством
предшественников, генерируемых с
помощью потери сигналов латеральной
ингибиции , у Notch, Delta или Su(H)
мутантов [69].
Генетический анализ взаимодействий
между латеральной ингибицией и
передачей сигналов Wingless указывает на
функцию Notch, отличную от ее роли в
латеральной ингибиции, которая
супрессирует развитие нейральных и
мышечных предшественников. Эта
функция м.б. антогонистичной для Wingless
сигналов в отношении развития
предшественников ([75••]; K Brennan, A Martinez Arias, unpublished observations). How Wingless signalling blocks this repression will require further study
[36••] . MAPK
белки и передача сигналов Wnt
Некоторые из внутриклеточных
компоенентов пути передачи
сигналов Wnt также участвуют во
множественных процессах (Fig. 1).Dishevelled
способен содействовать активации MAPK
белков подкласса JNK [76•][77•]
независимо от его влияния на
накопление β-catenin [77•].
Возмножно, что эта активность Dishevelled
не связана с каноничским путем
передачи сигналов Wnt [33••][34••][35••].
У C. elegans ранних эмбрионов,
эффективная передача сигналов Wnt
через POP-1 нуждается в активности MAPK
модуля, состоящего из MAPK-подобного
белка, кодируемого геном lit-1,[13][35••]
и активирующей MAP3K , кодируемой
геном Mom-4[35••].
Функция этого модуля находится в
связи с β-catenin, чтобы
супрессировать эффекты
репрессивных факторов POP-1/TCF и
таким образом участвовать в
сигнализации Wnt. Сходный MAPK
модуль существует у позвоночных в
форме TAK-1 (Mom-4) и NLK (Lit-1) [34••][35••]. TAK-1
киназа является членом семейства MAP3K ,
которое активируется с помощью TGF-β
и способно активировать членов JNK/p38
семейства MAPKs [78].
Так как TAK-1 может активровать
как JNK так и NLK/Lit-1 протеин киназу, то
возникает возможность, что эффекты Dishevelled
на JNK м.б.результатом способности
Dishevelled активировать вышестоящую
киназу TAK-1 и, тем самым NLK/Lit-1.
Если это так, то Dishevelled должен
участвовать в передаче сигналов Wnt
двумя путями: ингибируя Axin-зависимый
targeting of β-catenin для деградации и
участвуя в активации TAK-1 или NLK.
Недавно было показано, что NLK/Lit-1
киназа фосфорилирует TCF , если она
комплексуется с β-catenin, и
ингибирует его способность
связываться с ДНК [33••][35••].
Это наблюдение объясняет
потребность C. elegans как в Lit-1так
и в β-catenin чтобы супрессировать
активность TCF protein POP-1 во время
раннего предопределения судьбы
клеток вдоль передне-задней оси [13][15][16]. Wnt signalling revisited
У C. elegans, взаимодействие между
Lit-1/NLK и TCF необходимо для правильной
передачи сигналов Wnt [13][14••][34••].
Эффекты NLK в клетках
млекопитающих и у эмбрионов Xenopus[33••][35••],
указывают на то, что она возможно
играет некоторую роль и подтверждают,
что правильная передача сигналов Wnt
может зависеть от конвергенции двух
молекулярных событий:
стабилизации β-catenin и
активности Lit-1/NLK (Fig. 3).
Белок Lit-1/NLK существует также и у Drosophila[79]
и генетический анализ его
взаимодействия с передачей
сигналов Wnt дело будущего.
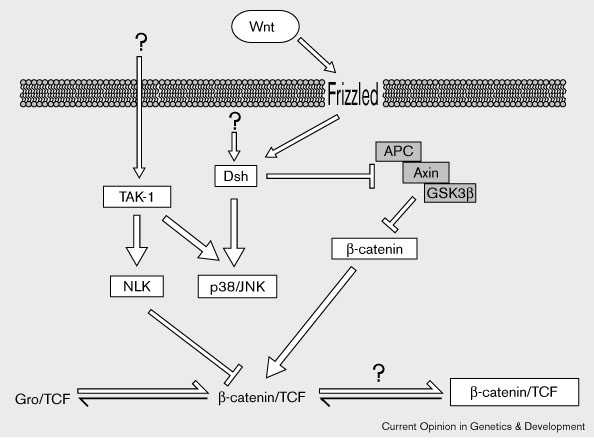
Figure 3
Интеграция активности NLK в
передачу сигналов Wnt . Активация
канонического пути передачи
сигналов Wnt вызывает
транслокацию β-catenin в ядро.
Ядерный β-catenin взаимодействует со
членами семейства TCF
транскрипционных регуляторов,
конкурирующих за свое связывание с
членами семейства Groucho (Gro) ко-репрессоров
(see main text and Fig. 1). NLK
модифицирует β-catenin/TCF комплекс
с помощью фосфорилирования TCF и
делает его неспособным связывать
ДНК. NLK активируется с помощью TAK-1,
который также активирует членов p38
и JNK семейства белков MAPK . Dishevelled,
помимо своего эффекта на
цитозольный пул β-catenin через GSK3β,
может активировать JNK. Неясно
как TAK-1 активируется in vivo,и как β-catenin/TCF комплекс может
становиться активатором (boxed) после
взаимодействия с NLK.
В терминах транскрипции может статься,
что основным состоянием в передаче
сигналов Wnt является репрессия,
обеспечиваемая с помощью TCF, вообще-то
через его взаимодействие с Groucho [80][81],
и что комбинация β-catenin и
активности Lit-1/NLK противодействует
этому взаимодействию с помощью
фосфорилирования TCF (Fig. 3).
В некоторых ситуациях , как у ранних
эмбрионов C. elegans, эта
комбинация будет гарантировать,
что TCF не вернется к репресси вному
состоянию. Это м.б. верно и для
некоторых промоторов, регулируемых Wnt
у Drosophila и позвоночных, оюнако ясно,
что у этих организмов имеются также
промоторы, которые нуждаются в
активаторах функции TCF [7].
Если комплекс TCF/β не связывается с
ДНК, когда фосфорилируется с помощью Lit-1/NLK [33••][35••],
то неясно как он становится
активатором. Вообще в этих случаях TCF
iне фосфорилируется с помощью NLK, или
является объектом дальнейших
модификаций. М.б., что влияние передачи
сигналов Wnt на транскрипцию
детерминируется балансом между
количеством ядерного β-catenin, TCF, и
активности Lit-1/NLK.
Как
упоминалось выше MAP3K кодируется Mom-4/TAK-1
геном, который является важным
элементом в передаче сигналов Wnt у C. elegans[34••].
Так как TAK-1 регулирует
активность NLK, то важно знать как
сам TAK1 регулируется внешними
сигналами и возникает. Одна из
возможностей, что TAK-1 активируется
членами TGF-β семейства сигнальных
молекул [78].
Возможно также, что Dishevelled может
способствовать активации TAK1 . В этой
связи важно, что белок Notch играет роль
в модуляции сигнализации JNK
независимо от его влияния на Su(H) [82•].
Боле того, делеция Dishevelled-связывающего
домена у Notch обусловливает строгое
подавление передачи сигналов с
помощью JNK [82•].
Возможно, что взаимодействия Notch с Dishevelled [72]
и Wingless [36••]
могут модулировать передачу сигналов
через TAK-1/NLK модуль и, таким
способом, влиять на конечный
результат передачи сигналов Wnt.
представляет собой информационную систему о ген-регуляторных путях, об элементах путей сигнальной трансдукции.