У позвоночных имеется два класса фоторецепторов: палочки и колбочки . Колбочки функционируют в широкой световой (дневной свет) амплитуде и ответственны за цветовое зрение. Палочки являются сенсорами слабого рассеянного света и не различают цвет. Зрение человека использует колбочки трех типов —blue, green and red—, а палочки только одного. Сетчатка человека содержит также центрально расположенную, богатую колбачками, область, называемую fovea, находящуюся в центре желтого пятна внутри сетчатки, называемой macula. fovea является областью наивысшей остроты зрения в сетчатке. Мыши и крысы обладают почти целиком опосродованным палочками зрением. Примерно 75% всех клеток сетчатки мыши являются фоторецепторами и 97% этих клеток являются палочками . Мыши имеют два варианта колбочек — с чувствительностью к средним (M) и коротким (S) длинам волн—, они не имеют fovea или macula
Каскад фототрансдукции начинается с улавливания света 11-cis-retinal, хромофорами, связанными с помощью опсиновых белков — родопсина в палочковых фоторецепторах и конусных опсинов в колбочковых фоторецепторах. Белки, которые передают фототрансдукцию располагаются в elaborate и высоко специализированных мембранных структурах,формирующих наружный сегмент каждого фоторецептора. Фоторецепторынй наружный сегмент является довольно ломким, дегенерирует в ответ на большинство средовых и/или генетических нарушений и вызывает слепоту.
Pellikka, M. et al. Crumbs, the Drosophila homologue of human CRB1/RP12, is essential for photoreceptor morphogenesis. Nature 2002 February 17 ( ) | |
Izaddoost, S. et al. Drosophila Crumbs is a positional cue in photoreceptor adherens junctions and rhabdomeres. Nature 2002 February 17 ( ) |
Тысячи людей слепнут из-за мутаций ретинальной дистрофии в — который кодирует белок, гомологичный регулятору Drosophila epithelial polarity, названному . Pellikka et al. и Izaddoost et al. сообщают, что Crumbs играет две специфические роли, отвечает за архитектуру photoreceptor cells (PRCs), которая формируется после ранней полризации клеток.
Обе группы использовали , чтобы удалить Crumbs из развивающейся сетчатки Drosophila. Кроме того его избыточно экспрессировали в PRCs мух. Ранее было показано, что Crumbs организует zonula adherens (ZA) — диск 'клея', который окружает каждую эпителиальную клетку и соединяет клетки вместе в один слой. Далее было установлено, что Crumbs поддерживает ZA интеграцию PRCs во время быстрой экспансии рабдомеров, органелл, которые упакованы фоточувствительными мембранами. Во время дифференцировки PRC рабдомеры (называемые наружным сегментом у позвоночных) драматически удлинняются и поддерживаются с помощью структур, называемых stalks (или внутренними сегментами у позвоночных). Потеря Crumbs из PRCs вызывает аномальное расположение ZA и сильное укорочение рабдомеров (Рис.), но не обнаруживается достоверных нарушений полярности эпителия. Izaddoost et al. показали, что при избыточной экспрессии внутриклеточных доменов как Crumbs так и CRB1 в Drosophila PRCs происходит сильное смещение ZA. Это указывает на то, что Crumbs играет роль в образовании ZA, которая отличается от его роли в обеспечении эпителиальной полярности, которая нуждается в др. структурных доменах белка и использует др. белковые партнеры.
Pellikka et al. сравнивали развивающиеся PRCs в сетчатке мух и мышей и нашли, что как только образуются мембраны отдельно для рабдомера (наружный сегмент) и ножки (внтуренний сегмент), то Crumbs и CRB1 становятся ограниченными ножкой и внутренним сегментом, соответственно. Показано также участие Crumbs в элонгации рабдомеров. Pellikka et al. показали, что внеклеточный домен Crumbs играет роль в формировании ножки также. Во-первых, они показали, что потеря Crumbs укорачивает ножки у мух, тогда как избыточная экспрессия его внеклеточного домена удлинняет их. Они установили также, что физически ассоциирует с Crumbs и вносит вклад в элонгацию ножки, возможно, в результате негативно регулируемого эндоцитоза. Все это указывает на то, что в развивающихся PRCs, Crumbs связывается посредством внутриклеточного домена с белковыми партнерами, чтобы предопределить позицию ZA. Как только ZA оказывается на месте, он взаимодействует с различными белковыми партнерами на внутренней и наружной поверхности клеток, чтобы поставлять мембраны в увеличивающуюся ножку.
Хотя глаза мух и человека совершенно отличны по своей морфологии и фототрансдукционному каскаду, однако имеются определенное сходство в морфогенезе фоторецепторов.
Наследственные болезни фоторецепторов можно разделить на наследственные макулярные дистрофии (inherited macular dystrophies, cone–rod dystrophies (CRDs)) и rod–cone dystrophies (называемых retinitis pigmentosa, RP). CRDs характеризуются потерей опосредуемого колбочками зрения в первой декаде жизни или позднее с последующей потерей опосредуемого палочками зрения. Напротив, RP характеризуется первоначальной потерей функции палочек, сопровождаемой потерей колбочками обеспечиваемого зрения. Большинство известных RP генов, также как и генов, ассоциированных с CRD, кодируют белки фоторецепторного наружного сегмента (см. RetNet Web site: ). Многие из этих белков необходимы для фототрансдукции или структуры наружного сегмента. Вопрос почему мутации генов, специфичных для палочек, обусловливают заболевания колбочек при RP остается неясным.
Выяснение причин глазных болезней с полигенной и мультифакторной основой, таких как зависимая от возраста макулярная дегенерация ( age-related macular degeneration (AMD)), возможно при изучении моногенных болезней. Например, двугенная форма RP возникает у пациентов, которые являются двойными гетерозиготами по мутациям несцепленных специфичных для фоторецепторов генов, кодирующих ROM1 и peripherin/RDS. Мутации в peripherin/RDS ответственны за доминантный моногенный RP. Анализе ABCR гена (кодирующего член семейства ABC transmembrane transporter), показал, что его дефекты ответственны за рецессивную наследственную макулярную дегенерацию при болезни Stargardt. Делеции и аминокислотные замены обнаружены в одиночном аллеле ABCR у 16% пациентов AMD, однако с этим объяснением несогласны другие исследователи
Описание развития фоторецепторов
У мышей и крыс спецификация типов клеток сетчатки осуществляется между 9-м днем эмбрионального развития (E9) и 7 постнатальным днем (P7). 7 типов клеток сетчатки возникают в эволюционно закрепленном порядке во время развития, хотя многие типы клеток продуцируются одновременно на каждой стадии развития. Колбочки генерируются рано в развитии, тогда как палочки возникают во время последних 2/3 ретиногенеза.
Установлено, что большинство, если не все, клеток сетчатки генерируется из мультипотентных предшественников. Так палочковые фоторецепторы часто обнаруживаются в клонах других типов клеток: включая биполярные или амакриновый интернейроны и даже Мюллеровскую глию, указывая что палочки возникают из мультипотентных предшественников, у грызунов часто встречаются клоны,содержащие только палочки. Это связано с обилием генерируемых палочек во время маркировки клона, но может указывать и на то, что существует отдельный предшественник палочек.
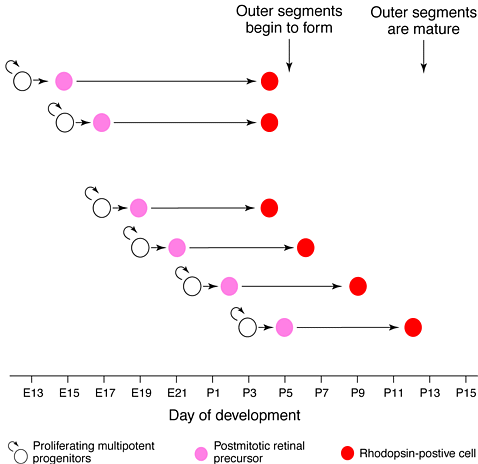
Имеются две разные фазы формирования палочек in vivo—ранняя и поздняя фазы Рис.1). Большинство предшественников палочек участвует в поздней фазе генеза палочек. Они появляются после E19 и это регулярно связано с началом экспрессии родопсина. Эти предшественники палочек начинают экспрессировать родопсин приблизительно спустя 5.5 - 6.5 дней после их терминального митоза. Напротив предшественники палочек ранней фазы появляются до E19, и эти клетки находятся в lag-фазе между последним митозом и началом экспрессии родопсина значительно дольше, от 8.5 до 12.5 days (Рис.1). Ранние предшественники палочек по-видимому инициируют экспрессию родопсина способом, который не скоррелирован во времени с терминальным митозом. Для этих клеток начало экспрессии родопсина, по-видимому, совпадает с поздно появляющимися клетками во время постнатального периода незадолго до формирования наружного сегмента. Экспрессия некоторых других специфичных для палочек генов практически мимикрирует экспрессию родапсина, начинаясь в основном в постнатальный период, примерно в P2.
Рис. 1. 2 отдельные фазы формирования палочек in vivo. Генез палочек делится на две фазы in vivo,раннюю и позднюю. Диаграмма показывает время lag-фазы от терминального митоза предшественника полочкек до начала экспрессии родопсина для кагорты, появляющейся с эмбрионального дня 15 (E15) и вплоть до постнатального дня 5 (P5). Для большинства предшественников палочек, появившихся после E19, время начала экспрессии родопсина тесно связанао со временем окончания клеточного цикла. Для этих клеток ступень, ведущая от ьерминального митоза к экспрессии родопсина занимает в среднем 5.5 - 6.5 дней. Эти клетки представляют позднюю фазу генеза палочек. Во время ранней фазы генеза палочек клетки, появившиеся до E19 обладают lag-фазой значительно более длинной, чем 6.5 дней, и начинают экспрессировать родопсин почти синхронно с клетками поздней фазы в раннем постнатальном периоде. Время инициации и окончания формирования наружного сегмента показано стрелками
Figure 1. Two distinct phases of rod development in vivo. Rod genesis is divided into two phases in vivo, an early phase and a late phase. This diagram depicts the time lags from the terminal mitosis of rod precursors to the onset of rhodopsin expression for cohorts born from embryonic day 15 (E15) through to postnatal day 5 (P5). For the majority of rod precursors, those born after E19, the timing of rhodopsin onset is tightly linked to the timing of cell-cycle withdrawal. For these cells, the steps leading from the terminal mitosis to rhodopsin expression take on average 5.5 to 6.5 days. These cells constitute the late phase of rod genesis. During the early phase of rod genesis, those cells born before E19 appear to display lags significantly longer than 6.5 days, and commence rhodopsin expression approximately synchronously with the late-phase cells in the early postnatal period. The initiation and completion times of formation of outer segments are marked by arrows.
Подобно палочкам ранней фазы колбочковые фоторецепторы появляются на ранних стадиях эмбриогенеза крыс не экспрессируют конусные описны вплоть до постнатального периода. S волновые колбочки появляются впервые на ст. P5. Пик S opsin-экспрессирующих клеток приходится на P10, сопровождается снижением этих singly-reactive клеток и последующим появлением колбочек, иммунореактивных как к S так и M колбочковым антителам. Эти `dual' колбочки затем, по-видимому, замещаются singly-immunoreactive M колбочками, указывая тем самым на прогресс дифференцировки колбочек от S колбочек, к дуальным колбочкам и ,наконец, к M колбочкам. Кинетика экспрессии разных опсинов относительно друг друга изучена у лягушек,золотых рыбок и кур, а их экспрессия относительно терминальных митозов также изучена на лягушках и рыбах.
Онтогенетическая пластичность после терминального митоза в предшествениках палочек
Предполагается,что мультипотентность предшественников сетчатки сохраняется до последнего клеточного деления или после него, когда клон состоит только из двух клеток с разными клеточными типами. Так было показано, что клетки, предназначенные стать палочками by virtue of their birthdate, были обработаны ciliary neurotrophic factor (CNTF) в культурах. Два дня спустя после терминальной S фазы предшественников палочек воздействие CNTF было достаточным для для блокирования экспрессиии родопсина и респецификации этих клеток предшественников к экспрессии по крайней мере трех маркеров биполяных интернейронов, указывая тем самым на изменение судьбы клеток. Это согласуется с моделью, согласно которой спецификация судьбы клеток палочек является прогрессивным процессом, который продолжается в постмитотических клетках, следовательно, отсутствует необратимое коммитирование и после терминального митоза (Рис.2). Клетки, которые уже экспрессируют обнаружимые уровни родопсина, резистентны к ингибирующему действию CNTF. Напротив, введение CNTF во взрослую сетчатку in vivo увеличивает выживаемость фоторецепторов в моделях дегенерации фоторецепторов.
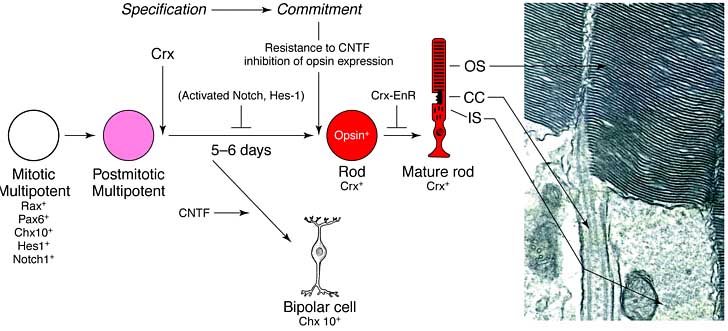
Рис. 2. Модель развития палочкового фоторецептора. Клональный анализ у многих видов показал, что палочки генерируются мультипотентными предшественниками. lag-период между последним митозом и синтезом опсина составляет 5.5–6.5 дня (см. Рис.1). Некоторые транскрипционные факторы экспрессируются в пуле митотически активных предшественников, продуцирующих палочки, включая три гена, кодирующих paired-типа гомеодоменовые белки, Rax, Pax6 and Chx10, негативно действующий basic helix–loop–helix (bHLH), HES-1 а также трансмембранный рецептор Notch 1 (подробнее в тексте). Некоторое время между завершением последнего митоза и экспрессией опсина, экспрессия всех этих генов затушена в клетках, которые станут палочками. Crx, другой paired-типа гомедоменовый ген, экспрессируется вскоре после предопределения судьбы клеток в направлении формирования палочек. Если активированный Notch1 аллель вносится в митотические клетки- предшественники палочекin vivo, то дифференцировка в зрелые палочки блокируется, а у мышей, гетерозиготных по HES-1-нуллевому аллелю, экспрессия родопсина появляется преждевременно в субнаборе палочек. Если ciliary neurotrophic factor (CNTF) добавляется к митотическим клеткам или постмитотическим клеткам, которые детерминированы экспрессировать опсин, но еще не начали его синтезировать, то экспрессия описна. У коыс, даже такие постмитотические клетки, предназначенные стать палочками, не комиттированы к судьбе палочек, так как часть этих клеток может экспрессировать, по крайней мере, три маркера детерминированныъх биполярных клеток. Если клетки экспрессируют обнаружимые количества опсина, то они резистентны к CNTF. Предполагается, что спецификация судьбы палочек происходит в течение нескольких дней, и начинается с информации, поступающей от митотических мультипотентных клеток их постмитотическим дочерним клеткам. Эта информация может одаривать клетки способностью становиться палочками в ответ на средовые сигналы. Эти сигналы могут быть частью процесса, который ингибирует экспрессию репертуара транскрипционных факторов, экспрессирующихся у мультипотентных предшественников и это ведет к синтезу Crx. Так и CNTF резистентность находится в соответствии с синтезом опсина, ступень коммитации, на которой клетки теряют пластичность и способны становиться биполярными нейронами, может происходить непосредственно перед началом синтеза опсина. Введение предполгаемого доминантного –негативного аллеля of Crx, в котором ДНК-связывающий домен Crx слит с репрессорным доменом Drosophila engrailed гена (Crx-EnR), ведет к отсуствию образования наружного сегмента и концов палочек. Справа: электронная микрофотография части палочковой фоторецепторной клетки . Обозначения: CC, connecting cilium; IS, inner segment; OS, outer segment.
Figure 2. A model of rod photoreceptor development. Lineage analyses in multiple species have shown that rods are generated by multipotent progenitors). The lag time between becoming postmitotic and synthesizing opsin is 5.5–6.5 days (см. Рис.1). Several transcription factors are expressed in the pool of mitotic progenitors that produce rods, including three genes that encode paired-type homeodomain proteins, Rax, Pax6 and Chx10, a negatively acting basic helix–loop–helix (bHLH), HES-1 and also the transmembrane receptor Notch 1 (see text for further details). Some time between becoming postmitotic and expressing opsin, expression of all of these genes is extinguished in cells that will become rods. Crx, another paired-type homeodomain gene, is expressed soon after a cell fated to become a rod is born. If an activated Notch1 allele is introduced into mitotic rod progenitor cells in vivo, differentiation into mature rods is blocked, and, in mice heterozygous for a HES-1-null allele, rhodopsin expression occurs precociously in a subset of rods. If ciliary neurotrophic factor (CNTF) is added to mitotic cells, or to postmitotic cells that are fated to express opsin, but have not yet begun to do so, the expression of opsin is blocked. In rats, even these postmitotic cells destined to be rods are not committed to the rod fate as a fraction of these cells go on to express at least three markers of the bipolar cell fate. Once a cell has expressed detectable levels of opsin, it is resistant to CNTF. We speculate that the specification to the rod fate occurs over several days, perhaps beginning with information passed from the mitotic multipotent cell to its postmitotic daughter. This information might endow a cell with the ability to become a rod in response to environmental cues. These cues in turn might be part of the process that extinguishes the expression of the repertoire of transcription factors expressed in the multipotent progenitor and that results in the synthesis of Crx. As CNTF resistance is concomitant with opsin synthesis, the commitment step, whereby a cell is no longer plastic and able to become a bipolar neuron, might occur just before opsin is detectable. Introduction of a presumed dominant–negative allele of Crx, in which the DNA-binding domain of Crx is fused to the repressor domain of the Drosophila engrailed gene (Crx-EnR), leads to lack of formation of outer segments and rod termini. Right-hand side: transmission electron micrograph of part of a rod photoreceptor cell, courtesy of E. Morrow. Abbreviations: CC, connecting cilium; IS, inner segment; OS, outer segment.
In vitro, basic fibroblast growth factor (bFGF), epidermal growth factor (EGF), taurine, S-laminin, thyroid hormone, retinoic acid, docosahexaenoic acid, glial-cell-line-derived neurotrophic factor (GDNF), Sonic hedgehog и CNTF семейство цитокинов все участвуют в детерминации, дифференцировке и/или выживаемости фоторецепторов. Роль этих факторовin vivo остается почти полностью неизученной.
Конструкция наружного сегмента фоторецептора
Наружный сегмент фоторецептора представлен колодой дисковых мембран, он связан с внутренним сегментом клетки с помощью nonmotile connecting cilium (Рис.2). В развивающихся фоторецепторах наружный сегмент генерируется из зарождающейся соединительной ножки (connecting cilium) с помощью удивительного процесса роста. В фоторецепторах взрослых диски продолжают формироваться в основании наружного сегмента, тогда как апикальные диски перестают функционировать, сбрасываются и фагоцитируются лежащим поверх пигментным эпителием. ROM1 и peripherin/RDS белки (локализованные в краях диска) и опсиновые белки (расположенные по всему диску) ,по-видимому, обеспечивают структурную интеграцию в зрелых наружных сегментах. Гомозиготные мутации по генам, кодирующим или родопсин или периферин/RDS, обусловливают полное отсутствие образования наружного сегмента.
У Drosophila, родопсин кодируетсяninaE геном и экспрессируется в фоторецепторах R1–R6. У ninaE-нуллевых мутантов рабдомеры, структурные аналоги наружных сегментов у позвоночных, неспособны развиваться в R1–R6 фоторецепторах, они напоминают ситуацию у rhodopsin-null мышей. У ninaE-нулевых мух ninaE трансген под контролем хитшокового промотора подвергался различным температурным сдвигам во время развития. Тепловой шок во время обычного начала экспрессии родопсина обусловливает существенное и продолжительное восстановление структуры фоторецепторов и временное восстановление физиологии фоторецепторов. Однако экспрессия незадолго до или после этого критического периода неспособна восстанавливать фоторецепторы, указывая тем самым, что экспрессия родопсина во время дискретного временного интервала в развитии существенна для собственно морфогенеза рабдомеров. Этот результат согласуется с наблюдениями у крыс, у которых начало появления родопсина приходится на ограниченный промежуток времени формирования большинства палочек in vivo.
Транскрипционная регуляция развития фоторецепторов
Длительная задержка между терминальным митозом предшественников палочек и началом экспрессии родопсина и неспособность укоротить этот lag-период с помощью воздействия на эти клетки различных rod-promoting условий, согласуются с идеей клеточно-автономного ингибитора экспрессии родопсина, присутствующего в только что появившихся предшественниках палочек. Несколько кандидатов транскрипционных факторов идентифицировано у позвовночных с паттернами экспрессии, указывающими на прямое или косвенное ингибирование дифференцировки палочек. Например, два гомеобоксных гена Pax6 и chx10, экспрессируются в предшественниках сетчатки. Их экспрессия исключается из развивающихся палочек, но поддерживается во внутреннем ядерном слое клеток, принципиально биполянрых интернейронов(chx10) и амакринных интернейронов(Pax6). Третий paired-типа гомеобоксный ген Rax экспрессируется в ретинальных предшественниках и последовательно подавляется во всех клетках после дифференцировки.
Надежные данные, связанные с ингибированием дифференцировки палочек, получены для Notch/HES сигнального пути. Экспрессия конституитивно активной формы Notch в развивающихся палочковых фоторецепторах in vivo у грызунов или Xenopus блокирует дифференцировку фоторецепторов. Сходные результаты получены для basic helix–loop–helix (bHLH) репрессора транскрипции HES-1,предполагаемого нижестоящего эффектора активности Notch. Кроме того Notch, HES-1 и HES-5 все подавляются во время дифференцировки фоторецепторов у грызунов. Более того CRS-3, защищеные последовательности вышестоящей области описна телят содержат предполагаемый HES-связывающий сайт, CACNAG. После нокаута HES-1 гена экспрессия родопсина, по-видимому, ускоряется, так что преждевременная экспрессия родопсина обнаруживается в сетчатке эмбрионов мыши. Ранняя экспрессия родопсина не только вторично обусловливает нарушение расположения тканей сетчатки, ранняя экспрессия родопсина обнаруживается и в сетчатке гетерозигот, которая имеет нормальную архитектуру. Однако неясно ускоряется ли весь генез палочек или укорачивается lag период между терминальным митозом и началом экспрессии родопсина.
Два позитивно действующих транскрипционных фактора участвуют в экспрессии фоторецепторных генов: the bZIP белок Nrl (neural retina leucine zipper)и гомеобоксный белок Crx (cone–rod homeobox). Nrl был первым транскрипционным фактором, который связываетмся и трансактивирует регуляторные элементы проксимальной промоторной области родопсина (RPPR) in vitro. Nrl экспрессия, однако, не ограничена фоторецепторами во время развития или взрослой сетчаткой. Роль Nrl iв развитии фоторецепторов и экспрессии родопсина in vivo остается невыясненной.
Crx новый член otd/Otx семейства гомеобоксных генов и обнаруживает 85% гомологию с Drosophila OTD гомоеобоксным доменом. Общее сходство между Crx и Otx1 и Otx2 вне гомеодомена низкое, но две аминокислотные последовательности законсервированы: WSP мотив и OTX хвостовой мотив. Во время развития Crx обнаруживает двухфазную экспрессию. СначалаCrx транскрипты обнаруживаются в развивающихся колбочковых фоторецептрах на ст.E12.5 мыши, а затем в в развивающихся палочковых фотоорецепторах. Во-вторых, строгая активация Crx обнаруживается поперек дифференцирующегося наружного ядерного слоя на ст.P6, непосредственно перед большой волной экспресиии родопсина и сегментным морфогенезом. Crx преимущественно специфичен для фоторецепторов в ходе развития, но экспрессируется и во взрослых фоторецепторах. Crx, следовательно, представляет собой первый специфичный для фоторецепторов транскрипционный фактор и Crx является в настоящее время самым ранним специфичным для фоторецепторов геном. (Crx транскрипты обнаружены также в шишковидной железе (pineal gland), ткани эволюционно связанной с фоторецепторами 58.) Кроме того Crx может связываться и трансактивировать регуляторные элементы разных специфичных для фоторецепторов генов, включая гены, кодирующие родопсин, interphotoreceptor retinoid binding protein (IRBP), arrestin и β-фосфодиэстеразу. Crx (или иммунологически сходный белок) присутствует в экстракте ядер из сетчатки, который связывается с RPPR элементами gel-shift системе и может активировать транскрипцию с родопсиновой промоторной конструкции синергично с Nrl in vitro.
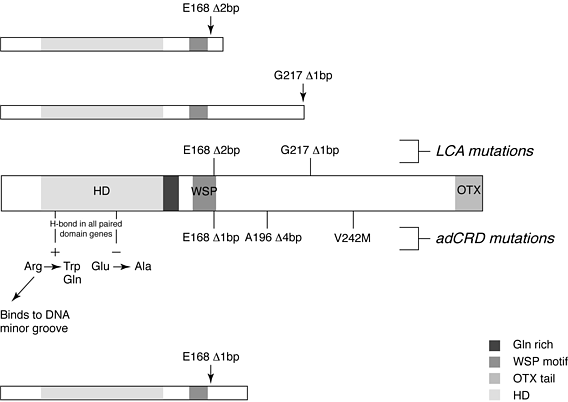
Рис. 3. Мутации в Crx, которые коррелируют с фоторецепторными болезнями. Показаны 4 области структуры Crx, которые законсервированы: HD—гомеодомен с 85–88% идентичностьтю с Otx1, Otx2 и Drosophila orthodenticle (otd): Gln rich—7 из 8 аминокислот являются глютамином; WSP— 7 из 13 аминокислот которого законсервированы у CRX, Otx1 и Otx2 имеющих последовательность WSP в центре; OTX хвост—область из 12 аминокислот, законервированная в генах otx позвоночных в виде одиночной или тандемных копий на С-конце. Мутации идентифицированы в семьях с cone–rod dystrophy 2 (adCRD2)и с Лберовским врожденным амаврозом (Leber's congenital amaurosis (LCA)). CRD2 аутосомно доминантное заболевание при котором обеспечиваемое как колбочками так и палочками зрение начинает ухудшаться между 10 и 20 годами. LCA аутосомно рецессивное заболевание, характеризуется слепотой с рожэдения. Мутации в законсервированной паре заряженных (charged) аминокислот в домене HD, аргинин и глютамин, ассоциируют с adCRD. Эти аминокислоты важны для формирования солевых мостиков, с аргининовой связью с малой бороздой мишени-ДНК. Мутации вблизи 3' конца домена WSP в E168 ассоциируют либо с LCA либо с adCRD2. При обоих заболеваниях формируются укороченные белки. В другой семье с LCA, мутация сдвига рамки считывания в G217 также обусловливает укорочение (как показано). missense мутация V242M и дополнительная мутация сдвига рамки A196/7 также ассоциирует с adCRD2. Обусловливает ли какая-либо из перечисленных мутаций болезнь в результате гаплонедостаточности или доминантно–негативного эффекта остается установить
Figure 3. Mutations in Crx that correlate with photoreceptor disease. Four regions of the structure of Crx that are conserved are shown: HD—homeodomain with 85–88% identity to Otx1, Otx2 and Drosophila orthodenticle (otd): Gln rich—seven of eight amino acids are glutamine; WSP—seven of 13 amino acids that are conserved among CRX, Otx1 and Otx2 having the sequence WSP in the centre; OTX tail—a region of 12 amino acids conserved among vertebrate otx genes as either single or tandem copies at the C-terminus. Mutations have been identified in families with cone–rod dystrophy 2 (adCRD2) and Leber's congenital amaurosis (LCA). CRD2 is an autosomal dominant disease in which both cone- and rod-mediated vision begin to deteriorate between 10 and 20 years of age. LCA is an autosomal recessive disorder characterized by blindness from birth. Mutations in a conserved pair of charged amino acids in the HD, arginine and glutamine, have been found in association with adCRD. These amino acids are important to the formation of a salt bridge, with the arginine binding to the minor groove of the target DNA. Mutations near the 3' end of the WSP domain at E168 are associated either with LCA or adCRD2. In both diseases, a truncated protein is formed, as depicted. In another family with LCA, a frameshift mutation at G217 also causes a truncation (as shown). A missense mutation, V242M, and an additional frameshift mutation, A196/7, have also been associated with adCRD2. Whether any of the above mutations cause disease due to haploinsufficiency or a dominant–negative effect remains to be determined. (Figure modified, with permission, from Ref. 65.)
Показано, что клетки сетчатки,экспрессирующие доминантно–негативную форму Crx из ретровирусного вектора мигрируют в наружный ядерный слой, подтверждая тем самым, что по крайней мере ограниченная спецификация судьбы фоторецепторных клеток происходит даже тогда, когда Crx транскрипционная активность нарушена. Однако эти презумптивные палочковые фоторецепторы имеют аберрантный фоторецепторный морфогенез, при котором не развиваются собственно наружные сегменты и окончания палочек.
CRX мутации при глазных болезнях человека
Мутации Crx обнаруживаются при двух глазных болезнях человека: Leber's congenital amaurosis (LCA и аутосоно доминантной cone–rod dystrophy 2 (adCRD2). LCA клинически гетерогенное заболевание, ранее принимавшееся за аутосомно рецессивное, которое обусловливает снижение или отсутствие фоторецепторной функции у детей. Предположительно обусловливающая болезнь микроделеция обнаружена в одном из аллелей пациента с LCA (Рис.3). Вторая мутация в Crx не найдена во второе аллеле этого пациента. (Однако регуляторная область не была изучена на присутсвие мутации.) Обе микроделеции обусловливают сдвиг рамки считывания так, что у предполагаемого белка должен отсутствовать С-конец, содержащий OTX хвост. Некоторые Crx мутации найдены у пациентов с adCRD2, включая две мутации внутри гомеодомена Crx(Рис.3). Ассоциация LCA и CRD2 через идентификацию болезнь-вызывающих мутаций в Crx указывает на интригующую взаимосвязь между развитем фоторецепторов и их дегенерацией. Имеется ли онтогенетический компонент в этиологии CRD2?
Crx может также играть роль в такой патологии человека как blue cone monochromacy. Обнаружена делеция в вышестоящей области пучка генов, кодирующих красные и зеленые опсины, при данной болезни. Эти делеции области в 579- п.н., которая имеет законсервованную в 37 п.н. стержневую последовательность, содержащуюю Otx гомеобокс-связывающий сайт; предполагается, что регуляция этих генов с помощью Crx нарушена.
Новый сигнальный путь от палочковых фоторецепторов к ганглиолярным клеткам сетчатки млекопитающих
Палочки очень чувствительны, они могут трансдуцировать абсорбцию одиночных фотонов и отвечают за зрение при низком уровне света, при моносвете. Колбочки менее чувствительны и обеспевают зрение в дневное время. И палочки и колбочки гиперполяризуются при увеличнии интенсивности света и тем самым высвобождают меньше своего тансмиттера, глютамата. Колбочки синапсируют с двумя типами биполярных клеток; один тип деполяризуется вследствие увеличения света и называется "ON bipolar" или 'depolarizing bipolar"; другой тип гиперполяризуется на свету и называется "OFF bipolar" или "hyperpolarizing bipolar" Разные реакции ON и OFF биполярных клеток предопределяются их глютаматовыми рецепторами: в OFF биполярных клетках глютамат вызывает приток натрия с помощью ионотропных рецепторов; в ON биполярных клетках метаботропные рецепторы связыают глютамат, чтобы закрыть катинные канальцы. ON биполярные клетки возбуждают ON ганглиолярные клетки, а OFF биполярные клетки возбуждают OFF ганглиолярные клетки. В результате временное увеличение интенсивности света передается в головной мозг с помощью быстрых burst of spikes с ON ганглиолярных клеток, тогда как временное снижениеинтенсивности кодируется в основном burst of spikes с OFF ганглиолярных клеток.
Палочки млекоптающих срединяются только с оджним типом биполярных клеток rod-depolarising bipolar (RDB). При увеличении интенсивности света эти нейроны возбуждают AII амакринные клетки, которые соединяются с двумя классами конусных биполярных клеток. Они возбуждают cone-depolarizing bipolar (CDB) клетки посредством электрических соединений и ингибируют cone-hyperpolarized bipolar (CHB) клетки посредством глицинэргических синапсов. Таким образом, параллельные ON и OFF пути возникают снова.
Предполагалось, что сигналы от палочек могут передаваться и другими путями, а именно через щелевые соединения к колбочкам и от них к On и OFF конусным биполярным клеткам. Электрические связи между палочками и колбочками известны.
Исследования на кроликах показали, что сигналы от палочек могут достигать ганглиолярных клеток сетчатки, минуя палочковые биполярные клетки.Этот путь осуществляется с помощью электрических щелевых соединений в результате чего сигналы от палочек прямо поступают в колбочки и конусные биполярные клетки. В сетчатке мышей также имеет место путь от палочек, минующий палочковые биполярные клетки AII. Этот альтернативный путь не нуждается в колбочковых фоторецепторах.