Regulation of retinal ganglion cell production by Sonic hedgehog
Xiang-Mei Zhang and Xian-Jie Yang
Ahmad, I., Dooley, C. M. and Polk, D. L. I. (1997). Delta-1 is a regulator ofneurogenesis in the vertebrate retina. Dev. Biol. 185, 92-103.
Alexiades, M. R. and Cepko, C. L.
(1997). Subsets of retinal progenitorsdisplay temporally regulated and distinct biases in the fates of their progeny.Development
124, 1119-1131.Altshuler, D., Turner, D. and Cepko, C. (1991). Specification of cell type inthe vertebrate retina. In Development of the Visual System (ed. M. Lam andC. Shatz), pp. 37-58. Cambridge: MIT Press.Altshuler, D. and Cepko, C.
(1992). A temporally regulated, diffusible activity is required for rod photoreceptor development in vitro. Development114, 947-957.Altshuler, D., Lo Turco, J. J., Rush, J. and Cepko, C.
(1993). Taurine promotes the differentiation of a vertebrate retinal cell type in vitro.Development 119, 1317-1328.Austin, C. P., Feldman, D. E., Ida, J. A., Jr and Cepko, C. L.
(1995).Vertebrate retinal ganglion cells are selected from competent progenitors bythe action of Notch. Development 121, 3637-3650.Baker, N. E., Yu, S. and Han, D.
(1996). Evolution of proneural atonal expression during distinct regulatory phases in the developing Drosophilaeye. Curr. Biol. 6, 1290-1301.Bao, Z. Z. and Cepko, C. L.
(1997). The expression and function of Notchpathway genes in the developing rat eye. J. Neurosci. 17, 1425-1434.Belecky-Adams, T., Cook, B. and Adler, R.
(1996). Correlations between terminal mitosis and differentiated fate of retinal precursor cells in vivo and in vitro: analysis with the ‘window- labeling’ technique. Dev. Biol. 178, 304- 315.Belliveau, M. J. and Cepko, C. L.
(1999). Extrinsic and intrinsic factors control the genesis of amacrine and cone cells in the rat retina. Development126, 555-566.Belloni, E., Muenke, M., Roessler, E., Traverso, G., Siegel-Bartelt, J., Frumkin, A., Mitchell, H. F., Donis-Keller, H., Helms, C., Hing, A. V. etal.
(1996). Identification of Sonic hedgehog as a candidate gene responsible for holoprosencephaly. Nat Genet 14, 353-356.Brown, N. L., Sattler, C. A., Paddock, S. W. and Carroll, S. B.
(1995). Hairy and emc negatively regulate morphogenetic furrow progression in the Drosophila eye. Cell 80, 879-887.Brown, N. L., Kanekar, S., Vetter, M. L., Tucker, P. K., Gemza, D. L. and Glaser, T.
(1998). Math5 encodes a murine basic helix-loop-helix transcription factor expressed during early stages of retinal neurogenesis.Development 125, 4821-4233.Cepko, C. L.
(1996). The patterning and onset of opsin expression in vertebrate retinae. Curr. Opin. Neurobiol. 6, 542-546.Cepko, C. L. (1999). The roles of intrinsic and extrinsic cues and bHLH genes in the determination of retinal cell fates. Curr. Opin. Neurobiol. 9, 37-46.Cepko, C. L., Austin, C. P., Yang, X., Alexiades, M. and Ezzeddine, D.
(1996). Cell fate determination in the vertebrate retina. Proc. Natl. Acad. Sci. USA 93, 589-595.Chen, S., Wang, Q. L., Nie, Z., Sun, H., Lennon, G., Copeland, N. G., Gilbert, D. J., Jenkins, N. A. and Zack, D. J.
(1997). Crx, a novel Otx-likepaired-homeodomain protein, binds to and transactivates photoreceptor cell-specific genes. Neuron 19, 1017-1030.Chiang, C., Litingtung, Y., Lee, E., Young, K. E., Corden, J. L., Westphal, H. and Beachy, P. A.
(1996). Cyclopia and defective axial patterning in mice lacking Sonic hedgehog gene function. Nature 383, 407-413.Dahmane, N. and Ruiz i Altaba, A.
(1999). Sonic hedgehog regulates the growth and patterning of the cerebellum. Development 126, 3089-3100.Dominguez, M.
(1999). Dual role for Hedgehog in the regulation of the proneural gene atonal during ommatidia development. Development 126, 2345-2353.Dominguez, M. and Hafen, E.
(1997). Hedgehog directly controls initiation and propagation of retinal differentiation in the Drosophila eye. Genes Dev 11, 3254-3264.Dorsky, R. I., Rapaport, D. H. and Harris, W. A.
(1995). Xotch inhibits cell differentiation in the Xenopus retina. Neuron 14, 487-496.Dorsky, R. I., Chang, W. S., Rapaport, D. H. and Harris, W. A.
(1997). Regulation of neuronal diversity in the Xenopus retina by Delta signalling. Nature 385, 67-70.Dowling, J. E.
(1987). The Retina - An Approachable part of the Brain. Cambridge, MA: Harvard University Press.Edlund, T. and Jessell, T. M.
(1999). Progression from extrinsic to intrinsic signaling in cell fate specification: a view from the nervous system. Cell 96, 211-224.Ekker, S. C., Ungar, A. R., Greenstein, P., von Kessler, D. P., Porter, J. A., Moon, R. T. and Beachy, P. A.
(1995). Patterning activities of vertebratehedgehog proteins in the developing eye and brain. Curr. Biol. 5, 944-955.Ericson, J., Morton, S., Kawakami, A., Roelink, H. and Jessell, T. M.
(1996). Two critical periods of Sonic Hedgehog signaling required for thespecification of motor neuron identity. Cell 87, 661-673.Ericson, J., Rashbass, P., Schedl, A., Brenner-Morton, S., Kawakami, A., van Heyningen, V., Jessell, T. M. and Briscoe, J.
(1997). Pax6 controls progenitor cell identity and neuronal fate in response to graded Shh signaling. Cell 90, 169-180.Ezzeddine, Z. D., Yang, X., DeChiara, T., Yancopoulos, G. and Cepko, C. L.
(1997). Postmitotic cells fated to become rod photoreceptors can be respecified by CNTF treatment of the retina. Development 124, 1055-1067.Fekete, D. M., Perez-Miguelsanz, J., Ryder, E. F. and Cepko, C. L.
(1994). Clonal analysis in the chicken retina reveals tangential dispersion of clonally related cells. Dev. Biol. 166, 666-682.Fuhrmann, S., Kirsch, M. and Hofmann, H. D.
(1995). Ciliary neurotrophic factor promotes chick photoreceptor development in vitro. Development121, 2695-2706.Furukawa, T., Morrow, E. M. and Cepko, C. L.
(1997). Crx, a novel otx-like homeobox gene, shows photoreceptor-specific expression and regulatesphotoreceptor differentiation. Cell 91, 531-541.Gavrieli, Y., Sherman, Y. and Ben-Sasson, S. A.
(1992). Identification ofp rogrammed cell death in situ via specific labeling of nuclear DNA fragmentation. J. Cell Biol. 119, 493-501.Goodrich, L. V. and Scott, M. P.
(1998). Hedgehog and patched in neural development and disease. Neuron 21, 1243-1257.Greenwood, S. and Struhl, G.
(1999). Progression of the morphogenetic furrow in the Drosophila eye: the roles of Hedgehog, Decapentaplegic and the Raf pathway. Development 126, 5795-5808.Guillemot, F. and Cepko, C. L.
(1992). Retinal fate and ganglion cell differentiation are potentiated by acidic FGF in an in vitro assay of early retinal development. Development 114, 743-754.Hallonet, M., Hollemann, T., Pieler, T. and Gruss, P.
(1999). Vax1, a novelhomeobox-containing gene, directs development of the basal forebrain andvisual system. Genes Dev. 13, 3106-3114.Hamburger, V. and Hamilton, H.
(1951). A series of normal stages in the development of the chick embryo. J. Morphol. 88, 49-92.Hammerschmidt, M., Brook, A. and McMahon, A. P.
(1997). The world according to hedgehog. Trends Genet. 13, 14-21.Harris, W. A.
(1997). Cellular diversification in the vertebrate retina. Curr.Opin. Genet. Dev. 7, 651-658.Heberlein, U. and Moses, K.
(1995). Mechanisms of Drosophila retinal morphogenesis: the virtues of being progressive. Cell 81, 987-990.Henrique, D., Hirsinger, E., Adam, J., Le Roux, I., Pourquie, O., Ish-Horowicz, D. and Lewis, J.
(1997). Maintenance of neuroepithelial progenitor cells by Delta-Notch signalling in the embryonic chick retina. Curr. Biol. 7, 661-670.Hicks, D. and Courtois, Y.
(1992). Fibroblast growth factor stimulates photoreceptor differentiation in vitro. J. Neurosci. 12, 2022-2033.Holt, C. E., Bertsch, T. W., Ellis, H. M. and Harris, W. A.
(1988). Cellular determination in the Xenopus retina is independent of lineage and birth date. Neuron 1, 15-26.Hu, M. and Easter, S. S.
(1999). Retinal neurogenesis: the formation of the initial central patch of postmitotic cells. Dev. Biol. 207, 309-321.Hughes, S. H., Greenhouse, J. J., Petropoulos, C. J. and Sutrave, P.
(1987). Adaptor plasmids simplify the insertion of foreign DNA into helper-independent retroviral vectors. J. Virol. 61, 3004-3012.Jarman, A. P., Grell, E. H., Ackerman, L., Jan, L. Y. and Jan, Y. N.
(1994). Atonal is the proneural gene for Drosophila photoreceptors. Nature 369,398-400.
Jasoni, C. L., Walker, M. B., Morris, M. D. and Reh, T. A.
(1994). A chicken achaete-scute homolog (CASH-1) is expressed in a temporally and spatially discrete manner in the developing nervous system. Development 120, 769- 783.Jensen, A. M. and Wallace, V. A.
(1997). Expression of Sonic hedgehog and its putative role as a precursor cell mitogen in the developing mouse retina. Development 124, 363-371.Kanekar, S., Perron, M., Dorsky, R., Harris, W. A., Jan, L. Y., Jan, Y. N. and Vetter, M. L.
(1997). Xath5 participates in a network of bHLH genes in the developing Xenopus retina. Neuron 19, 981-994.Kelley, M. W., Turner, J. K. and Reh, T. A.
(1994). Retinoic acid promotes differentiation of photoreceptors in vitro. Development 120, 2091-2102.Koshiba-Takeuchi, K., Takeuchi, J. K., Matsumoto, K., Momose, T., Uno, K., Hoepker, V., Ogura, K., Takahashi, N., Nakamura, H., Yasuda, K. and Ogura, T.
(2000). Tbx5 and the retinotectum projection. Science 287, 134-137.Levine, E. M., Roelink, H., Turner, J. and Reh, T. A.
(1997). Sonic hedgehog promotes rod photoreceptor differentiation in mammalian retinal cells in vitro. J Neurosci 17, 6277-6288.Lillien, L.
(1995). Changes in retinal cell fate induced by overexpression of EGF receptor. Nature 377, 158-162.Lillien, L.
(1998). Neural progenitors and stem cells: Mechanisms of progenitor heterogeneity. Curr. Opin. Neurobiol. 8, 37-44.Liu, W., Khare, S. L., Liang, X., Peters, M. A., Liu, X., Cepko, C. L. and Xiang, M.
(2000). All Brn3 genes can promote retinal ganglion cell differentiation in the chick. Development 127, 3237-3247.Macdonald, R., Barth, K. A., Xu, Q., Holder, N., Mikkola, I. and Wilson, S. W.
(1995). Midline signalling is required for Pax gene regulation and patterning of the eyes. Development 121, 3267-3278.Marigo, V., Davey, R. A., Zuo, Y., Cunningham, J. M. and Tabin, C. J.
(1996). Biochemical evidence that patched is the Hedgehog receptor. Nature38
4, 176-179.Masai, I., Stemple, D. L., Okamoto, H. and Wilson, S. W.
(2000). Midline signals regulate retinal neurogenesis in zebrafish. Neuron 27, 251-263.Masland, R.H. and Raviola, E.
(2000). Confronting complexity: strategies for understanding the microcircuitry of the retina. Annu. Rev. Neurosci. 23, 249-284.McCabe, K. L., Gunther, E. C. and Reh, T. A.
(1999). The development of the pattern of retinal ganglion cells in the chick retina: mechanisms that control differentiation. Development 126, 5713-5724.McFarlane, S., Zuber, M. E. and Holt, C. E.
(1998). A role for the fibroblast growth factor receptor in cell fate decisions in the developing vertebrate retina. Development 125, 3967-3975.McMahon, A. P.
(2000). More surprises in the Hedgehog signaling pathway. Cell 100, 185-188.Ming, J. E., Roessler, E. and Muenke, M.
(1998). Human developmental disorders and the Sonic hedgehog pathway. Mol. Med. Today 4, 343-349.Morgan, B. A. and Fekete, D. M.
(1996). Manipulating gene expression with replication-competent retroviruses. Methods Cell Biol. 51, 185-218.Morrow, E. M., Furukawa, T., Lee, J. E. and Cepko, C. L.
(1999). NeuroD regulates multiple functions in the developing neural retina in rodent. Development 126, 23-36.Morrow, E. M., Belliveau, M. J. and Cepko, C. L.
(1998). Two phases of rod photoreceptor differentiation during rat retinal development. J. Neurosci. 18, 3738-3748.Neumann, C. J. and Nuesslein-Volhard, C.
(2000). Patterning of the zebrafish retina by a wave of sonic hedgehog activity. Science 289, 2137- 2139.Perron, M., Opdecamp, K., Butler, K., Harris, W. A. and Bellefroid, E. J.
(1999). X-ngnr-1 and Xath3 promote ectopic expression of sensory neuron markers in the neurula ectoderm and have distinct inducing properties in the retina. Proc. Natl. Acad. Sci. USA 96, 14996-15001.Pignoni, F. and Zipursky, S. L.
(1997). Induction of Drosophila eye development by decapentaplegic. Development 124, 271-278.Prada, C., Medina, J. I., Lopez, R., Genis-Galvez, J. M. and Prada, F. A.
(1992). Development of retinal displaced ganglion cells in the chick: neurogenesis and morphogenesis. J. Neurosci. 12, 3781-3788.Reh, T. A. and Levine, E. M.
(1998). Multipotential stem cells and progenitors in the vertebrate retina. J. Neurobiol. 36, 206-220.Reh, T. A. and Tully, T.
(1986). Regulation of tyrosine hydroxylase-containing amacrine cell number in larval frog retina. Dev. Biol. 114, 463- 469.Riddle, R. D., Johnson, R. L., Laufer, E. and Tabin, C.
(1993). Sonic hedgehog mediates the polarizing activity of the ZPA. Cell 75, 1401-1416.Roessler, E., Belloni, E., Gaudenz, K., Jay, P., Berta, P., Scherer, S. W., Tsui, L. C. and Muenke, M.
(1996). Mutations in the human Sonic Hedgehog gene cause holoprosencephaly. Nat. Genet. 14, 357-360.Schulte, D., Furukawa, T., Peters, M. A., Kozak, C. A. and Cepko, C. L.
(1999). Misexpression of the Emx-related homeobox genes cVax and mVax2 ventralizes the retina and perturbs the retinotectal map. Neuron 24, 541-553.Snow, R. L. and Robson, J. A.
(1994). Ganglion cell neurogenesis, migration and early differentiation in the chick retina. Neuroscience 58, 399-409.Spence, S. G. and Robson, J. A.
(1989). An autoradiographic analysis of neurogenesis in the chick retina in vitro and in vivo. Neuroscience 32, 801- 812.Stenkamp, D. L., Frey, R. A., Prabhudesai, S. N. and Raymond, P. A.
(2000). Function for hedgehog genes in zebrafish retinal development. Dev. Biol. 220, 238-252.Stoker, A. W. and Bissell, M. J.
(1987). Quantitative immunocytochemical assay for infectious avian retroviruses. J. Gen. Virol. 68, 2481-2485.Tomlinson, A. and Ready, D. F.
(1987). Neuronal differentiation in the Drosophila ommatidium. Dev. Biol. 120, 366-376.Torelli, S., Sogos, V., Marzilli, M. A., D’Atri, M. and Gremo, F.
(1989). Developmental expression of intermediate filament proteins in the chick embryo retina: in vivo and in vitro comparison. Exp. Biol. 48, 187-196.Treisman, J. E. and Heberlein, U.
(1998). Eye development in Drosophila: formation of the eye field and control of differentiation. Curr. Top. Dev. Biol. 39, 119-158.Turner, D. L. and Cepko, C. L.
(1987). A common progenitor for neurons and glia persists in rat retina late in development. Nature 328, 131-136.Turner, D. L., Snyder, E. Y. and Cepko, C. L.
(1990). Lineage-independent determination of cell type in the embryonic mouse retina. Neuron 4, 833- 845.Waid, D. K. and McLoon, S. C.
(1995). Immediate differentiation of ganglion cells following mitosis in the developing retina. Neuron 14, 117-124.Waid, D. K. and McLoon, S. C.
(1998). Ganglion cells influence the fate of dividing retinal cells in culture. Development 125, 1059-1066.Wallace, V. A. and Raff, M. C.
(1999). A role for Sonic hedgehog in axon-to-astrocyte signalling in the rodent optic nerve. Development 126, 2901- 2909.Watanabe, T. and Raff, M. C.
(1990). Rod photoreceptor development in vitro: intrinsic properties of proliferating neuroepithelial cells change as development proceeds in the rat retina. Neuron 4, 461-467.Watanabe, T. and Raff, M. C.
(1992). Diffusible rod-promoting signals in the developing rat retina. Development 114, 899-906.Wechsler-Reya, R. J. and Scott, M. P.
(1999). Control of neuronal precursor proliferation in the cerebellum by Sonic Hedgehog [see comments]. Neuron 22, 103-114.Wetts, R. and Fraser, S. E.
(1988). Multipotent precursors can give rise to all major cell types of the frog retina. Science 239, 1142-1145.Wei, Y. and Allis, C.D.
(1998). Pictures in cell biology. Trends Cell Biol. 8, 266.White, N. M. and Jarman, A. P.
(2000). Drosophila atonal controls photoreceptor R8-specific properties and modulates both receptor tyrosine kinase and Hedgehog signalling. Development 127, 1681-1689.Wolff, T. and Ready, D.F.
(1993). Pattern formation in the Drsophila retina. In The Development of Drosophila melanogaster. (ed. M Bate and A. Martinez-Arias). pp. 1277-1326. Cold Spring Harbor: Cold Spring Harbor Laboratory Press.Yamada, T., Pfaff, S. L., Edlund, T. and Jessell, T. M.
(1993). Control of cell pattern in the neural tube: motor neuron induction by diffusible factors from notochord and floor plate. Cell 73, 673-686.Yan, R. T. and Wang, S. Z.
(1998). NeuroD induces photoreceptor cell overproduction in vivo and de novo generation in vitro. J. Neurobiol. 36, 485-496.Yang, X. J. and Cepko, C. L.
(1996). Flk-1, a receptor for vascular endothelial growth factor (VEGF), is expressed by retinal progenitor cells. J. Neurosci. 16, 6089-6099.Ye, W., Shimamura, K., Rubenstein, J. L., Hynes, M. A. and Rosenthal, A.
(1998). FGF and Shh signals control dopaminergic and serotonergic cell fate in the anterior neural plate. Cell 93, 755-766.Young, R. W.
(1985). Cell differentiation in the retina of the mouse. Anat. Rec. 212, 199-205.Yourey, P. A., Gohari, S., Su, J. L. and Alderson, R. F.
(2000). Vascular endothelial cell growth factors promote the in vitro development of rat photoreceptor cells. J. Neurosci. 20, 6781-6788.Zhang, X.-M. and Yang, X.-J.
(2001). Temporal and spatial effects of Sonic hedgehog signaling in chick eye morphogenesis. Dev. Biol., in press.Note added in proof
Gonzalez-Hoyuela, M., Barbas, J. A. and Rodriguez-Tebar, A.
(2001). The autoregulation of retinal ganglion cell number. Development 128, 117-124.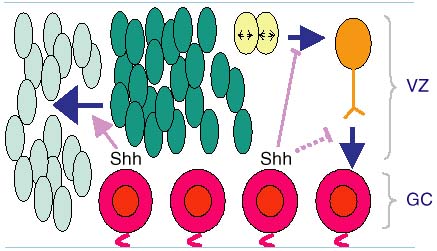
В сетчатке мышей на соотв. ст. развития наблюдается сходные паттерн экспрессии Shh и Ptc (Jensen and Wallace, 1997), а в сетчатке рыбок данио экспрессируются и shh и twhh в ганглиолярных клетках (Neumann and Nuesslein-Volhard, 2000).
| |
| Pea3 Expression Is Regulated by FGF Signaling In Developing Retina K.L.McCabe (klm@caltech.edu), C McGurie, T.A.Reh Dev. Dyn. - 2006.- V.235, No 2 - P. 327-335 |