Молчание генов
| ORIGINAL RESEARCH PAPERS Kirchmaier, A. & Rine, J. DNA replication-independent silencing in S. cerevisiae. Science 291, 646-650 (2001) | | | | Li, Y.-C., Cheng, T.-H. & Gartenberg, M. R. Establishment of transcriptional silencing in the absence of DNA replication. Science 291, 650-653 (2001) | | | | FURTHER READING Smith, J. S. & Boeke, J. D. Is S phase important for transcriptional silencing? Science 291, 608-609 (2001) | | | | WEB SITE Chromatin structure and function |
Между ранней S фазой и G2/M происходит сдвиг клеточного цикла, который позволяет сделать экспрессию генов молчащей. В ответ на это событие хроматин воспринимает измененную форму - гетерохроматина - которая предупреждает транскрипционную кухню (machinery) от нахождения своих генов-мишеней. Репликация ДНК подстегивает это переключение.
Анализировали у почкующихся дрожжей локус типа спаривания, который молчит во время образования гетерохроматина. нуждается в связывании некоторых белков с цис-действующими элементами, называемыми сайленсерами (silencers). Среди этих белков находится и origin-recognition complex (ORC), который участвует в инициации репликации ДНК (однако эта функция отделима от его роли в silencing). ORC затем рекрутирует Sir (silent information regulator) белок , а он, в свою очередь, облегчает включение дальнейших Sir белков в гетерохроматин.
Сравнивали образование гетерохроматина на хромосомном HMRa на нереплицирующейся, экстрахромосомной кольцевой ДНК (которая вырезается с помощью сайт-специфисеской рекомбинации)и не м. реплицироваться из-за отсутствия ORC-связывающего сайта. Для выяснения необходимости ORC в silencing, авт. сконструировали или Gal4 (Kirchmaier and Rine) или LexA (Li and colleagues) DNA-связывающие сайты во внехромосомном кольце. Для экспрессии Gal4-Sir1 или LexA-Sir1 слитых белков предоставляли все инградиенты, необходимые для silencing.
Выявлены незначительные отличия в silencing двух HMRa локусов, следовательно, репликация не нужна для инициации этого процесса. Подтверждено, что прохождение через S фазу обязательно для silencing.
Но если не репликация, то что вызывает переключение? Предполагается, что активность silencing компонентов - и вообще Sir белков - контролируется cell-cycle-зависимым образом, возможно, регулируя синтез кофакторов или ингибиторов деградации.
|
Gene silencing: double-stranded RNA mediated mRNA degradation and gene inactivation Wei T, Luo X.Y, Sanmuels V. Cell Res. V.11. N 3.P. 181-186. 2001 |
РНК участвует в двух типах homology-dependet gene silencing (HDGS): 1)Посттранскрипционном молчанием генов (PTGS), связанным с целенаправленной деградацией гомологичныхРНК в цитоплазме; 2) РНК-управляемым метилированием ДНК (RdDM), которое м.б. индуцировано с помощью РНК, получаемой с гомологичных ДНК последовательностей на уровне генома. Механизмы этих разных способов HDGS и характеристика участвующих РНК активно исследуются. Двунитчатая РНК (днРНК) играет двойную роль в молчании генов у растений, инициируя ступень РНК-деградации как в PNGS, так и в RdSM.
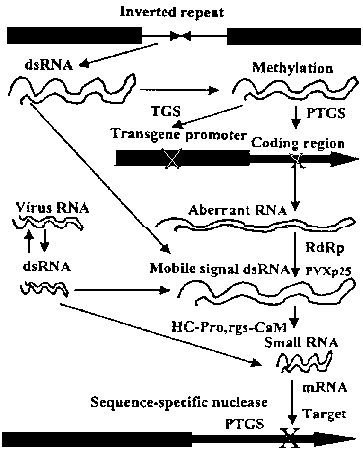 Модель dsRNA-обусловленного генного молчания. Показаны ступени, с вовлечением dsRNA, и ступени, которые затрагиваются вирусными супрессорами потстранскрипционного генного молчания и у разных PTGS мутантов. Транскрипционное генное молчание м.б. запущено непосредственно с помощью транскрипции последовательностей инвертированных посторов в ядре и с помощью метилирования гомологичных промоторных областей в геноме. dsRNA и др. аберрантные РНК формируются в ядре и вступают на путь PTGS. Индуцированное вирусом молчание генов обеспечивается вирусным RdRP, а индуцированное трансгенами молчание обеспечивается клеточной RdRP, что ведет к PTGS в цитоплазме. dsRNA из обоих этих источников м.б. отловлена с помощью предполагаемой dsRNA специфической рибонуклеазы, которая генерирует РНК в 21-25 нуклеотидов. PTGS м.б. индуцирован локально и затем распространипться с помощью мобильных сайленсинг сигналов. РС-Pro супрессирует молчание генов путем активирования эндогенного клеточного супрессора PTGS, rgs-CaM. PVX p25 супрессор PTGS предупреждает гнакопление мобильных сайленсинг сигналов путем взаимодействия с клеточной RdRP ветвью пути.
Модель dsRNA-обусловленного генного молчания. Показаны ступени, с вовлечением dsRNA, и ступени, которые затрагиваются вирусными супрессорами потстранскрипционного генного молчания и у разных PTGS мутантов. Транскрипционное генное молчание м.б. запущено непосредственно с помощью транскрипции последовательностей инвертированных посторов в ядре и с помощью метилирования гомологичных промоторных областей в геноме. dsRNA и др. аберрантные РНК формируются в ядре и вступают на путь PTGS. Индуцированное вирусом молчание генов обеспечивается вирусным RdRP, а индуцированное трансгенами молчание обеспечивается клеточной RdRP, что ведет к PTGS в цитоплазме. dsRNA из обоих этих источников м.б. отловлена с помощью предполагаемой dsRNA специфической рибонуклеазы, которая генерирует РНК в 21-25 нуклеотидов. PTGS м.б. индуцирован локально и затем распространипться с помощью мобильных сайленсинг сигналов. РС-Pro супрессирует молчание генов путем активирования эндогенного клеточного супрессора PTGS, rgs-CaM. PVX p25 супрессор PTGS предупреждает гнакопление мобильных сайленсинг сигналов путем взаимодействия с клеточной RdRP ветвью пути.
Модель dsRNA-обусловленного генного молчания. Показаны ступени, с вовлечением dsRNA, и ступени, которые затрагиваются вирусными супрессорами потстранскрипционного генного молчания и у разных PTGS мутантов. Транскрипционное генное молчание м.б. запущено непосредственно с помощью транскрипции последовательностей инвертированных посторов в ядре и с помощью метилирования гомологичных промоторных областей в геноме. dsRNA и др. аберрантные РНК формируются в ядре и вступают на путь PTGS. Индуцированное вирусом молчание генов обеспечивается вирусным RdRP, а индуцированное трансгенами молчание обеспечивается клеточной RdRP, что ведет к PTGS в цитоплазме. dsRNA из обоих этих источников м.б. отловлена с помощью предполагаемой dsRNA специфической рибонуклеазы, которая генерирует РНК в 21-25 нуклеотидов. PTGS м.б. индуцирован локально и затем распространипться с помощью мобильных сайленсинг сигналов. РС-Pro супрессирует молчание генов путем активирования эндогенного клеточного супрессора PTGS, rgs-CaM. PVX p25 супрессор PTGS предупреждает гнакопление мобильных сайленсинг сигналов путем взаимодействия с клеточной RdRP ветвью пути. Gene silencing mechanism mediated by dsRNA
Исследованияи на растениях было установлено, что молчание сопровождается накоплением малых РНК (21-25 нуклеотидов) как в смысловой, так и антисмысловой ориентации, которые гомологичны молчащему локусу, возникающих в результате сиквенс-специфической деградации РНК. Молчание м. запускаться локально и затем распространяться по организму растения с помощью мобильных silencing сигналов. Согласно модели сайленсинга происходит следующее: 1) синтез и амплификация днРНК; 2) расправление днРНК; и 3)нахождение мРНК после связывания с рибосомами. Клеточная РНК-управляемая РНК полимараза (RdRP) играет центральную роль в пострансляционном сайленсинге генов. Ее возможная функция - синтез cRNA с абератных РНК матриц, ведущий к образованию днРНК.
HC-Pro (helper component proteinase) супрессия постранскрипционного молчания не взаимодействует ни с продукцией, ни с движением сайленсинг сигнала, а предупреждает растения от реакции на этот сигнал, т.е. действует на ступень, предшествующую накоплению малых РНК. Идентифицирован calmodulin-родственный белок (rgs-CaM), который взаимодействует с HС-Pro. Этот rgsCaM белок супрессирует молчание генов и м.б. промежуточной субстанцией в супрессии посттранскрипционного молчания с помощью РС-Pro. Так как калмодулин и родственнные ему белки действуют связывая кальций и тем активируя белки-мишени, то РС-Pro супрессия возможно осуществляется за счет активации rgs-CaM и неизвестного белка-мишени. Белок PVX p25? повидимому, супрессирует посттранскрипционное молчание с помощью нахождения мобильного сайленсинг сигнала. Малые смысловые и антисмысловые РНК, связанные с молчанием, возникают в результате расщепления днРНК, а специфическая рибонуклеаза обеспечивает нахождение гомологичных мишеней РНК. Малые РНК накапливаются при молчании генов, индуцированных вирусом и трансгеном. Это указывает на конвергенцию этих двух процессов молчания до образования сиквенс-специфической рибонуклеазы.
Methylation and posttranscriptional gene silencing
Метилирование или кодирующих или транскрипбируемых областей трансгенов обнаруживается во многих случаях PTGS у растений. Возникновение и поддержание PTGS, по-видимому, нуждается в эпигенетических модификациях ДНК. Механизм RdDM неизвестен, но предполагается, что он связан с РНК-ДНК взаимодействием, базирующемся на помологии последовательностей. Минимальный размер мишени ДНК для RdDM 30 п.н., что позволяет предположить, что продукты дегардации РНК в 21-25 нуклеотидов днДНК ответственны за направление метилирования. Малые РНК м., по-видмому, вести ДНК метил трансферазу к немодифицированным гомологичным ДНК последовательностям.
RNA-directed DNA methylation (RdDM) представляет собой альтернативный способ индукции сиквенс специфического метилирования, наблюдаемого как при посттранскрипционном, так и транскрипционном молчении генов. RdDM вызывает плотное метилирование большинства симметричных и несимметричных цитозинов внутри области гомологии между индуцирующей РНК и мишенью ДНК. Метилирование последовательностей дупликаций, ассоциирванное с HDGS (homology-dependet gene silencing), связано с ДНК-ДНК спариванием и ДНК мишенями в 30 п.н., котоые м.б. модифицированы.
Cellular protein involved in posttranscriprional gene silencing
Хотя потребность в RdRP очевидна, природа субстрата для этого энзима неясна. RdRP, как полагают, использует предсуществующую lyHYR или аберантную смысловую РНК как матрицы для для синтеза антисмысловых РНК. Аберантная РНК - неправльно сплайсирована или закончена.
|
Post-transcriptional Gene Silencing By Double-stranded RNA Scott M. Hammond, Amy A. Caudy & Gregory J. Hannon Nature Reviews Genetics 2, 110 -119 (2001) |
| How does RNAi work? Delivering double-stranded RNA What does RNA interference do for me? |
RNA interference (RNAi) - это процесс, с помощью которого двунитчатая РНК специфически делает молчащей (silences) экспрессию гомологичных генов благодаря деградации их cognate мРНК. Сайленсинг (Silencing), следовательно, пост-транскрипционный феномен.
Генетическое молчание, которое наступает с помощью процесса RNAi, высоко специфично и базируется на последовательностях двунитчатой РНК.
Пост-транскрипционное молчение генов у растений и грибов механистически сходно с процессом RNAi у животных.
Деструкция мРНК обеспечивается мультикомпонентными нуклеазами, называемыми RNA-induced silencing complex, или RISC.
Внесенные двунитчатые РНК подвергаются процессингу на короткие РНК в 22 нуклеотида, которые инкорпорируются RISC. Здесь они действуют как РНК-наводчики ('guide RNAs'), сообщая нуклеазам специфические особенности мишеней.
Ряд молекул, которые участвуют в процессе RNAi, идентифицирован во многих модельных системах.
Предполагается, что биологической ролью RNAi является резистентность к вирусам, превращение транспозонов в молчащие и регуляция экспрессии эндогенных генов, в частнрости во время развития.
RNAi становится ценным экспреиментальным инструментом для исследования функции генов у Caenorhabditis elegans, Drosophila melanogaster и растений.
Когда механизм RNAi будет более понятен он может стать мощным технологическим средством для изучения широкого круга систем, включая и организмы млекопитающих и культивируемые клетки.
The Complexity of Gene Silencing
The Complexity of Gene Silencing Jim Kling
The Scientist 16[4]:31, Feb. 18, 2002
The Scientist 16[4]:31, Feb. 18, 2002
Researchers demonstrate the physical association between histone deacetylases and methyl-CpG binding proteins
1. E. Russo, "Hot
papers in acetylation," and "Hot papers in deacetylation," .
2. B.A. Maher, "Researchers focus on histone code," .
3.Y. Zhang et al., "Analysis of the NuRD subunits reveals a histone deacetylase core complex and a connection with DNA methylation," Genes & Development, 13:1924-35, 1999. (Cited in 117 papers).
4.S.K. Bhattacharya et al., "A mammalian protein with specific demethylase activity for mCpG DNA," Nature, 397:579-83, 1999.
5. B. Hendrich et al., "Closely related proteins MBD2 and MBD3 play distinctive but interacting roles in mouse development," Genes & Development, 15:710-23, March 15, 2001.
Jim Kling interviewed University of Edinburgh geneticist Adrian Bird and Paul A. Wade, assistant professor in the department of pathology and laboratory medicine at Emory University School of Medicine in Atlanta.
2. B.A. Maher, "Researchers focus on histone code," .
3.Y. Zhang et al., "Analysis of the NuRD subunits reveals a histone deacetylase core complex and a connection with DNA methylation," Genes & Development, 13:1924-35, 1999. (Cited in 117 papers).
4.S.K. Bhattacharya et al., "A mammalian protein with specific demethylase activity for mCpG DNA," Nature, 397:579-83, 1999.
5. B. Hendrich et al., "Closely related proteins MBD2 and MBD3 play distinctive but interacting roles in mouse development," Genes & Development, 15:710-23, March 15, 2001.



