В результате сомитогенеза несегментированная параксиальная мезодерма подразделяется на парные эпителиальные сферы, называемые сомитами. Путь передачи сигналов Notch является важным для регуляции формирования передне-заднего паттерна сомитов у позвоночных. Один из компонентов сигнального пути Notch ген fringle у дрозофилы кодирует секретируемую сигнальную молекулу, необходимую для для активации Notch. У мышей, гомозиготных по мутации гена lunatic fringle (Lfng), обнаруживаются дефекты формирования сомитов. Сомиты у мутантных эмбрионов нерегулярны по размерам и форме и их передне-задний паттерн нарушен. Показано, что в пресомитной мезодерме у мутантных эмбрионов резко демаркированные домены экспрессии некоторых компонентов сигнального пути Notch замещены градиентами генной экспрессии (Zhang, Gridley T., 1998).
Ген lunatic fringle кодирует секретируемый фактор, который функционирует как специфичная для границ сигнальная молекула, необходимая для для локализации сигнальной активности белка Notch. Мутантные по lunatic fringle мыши неспособны формировать границы между отдельными сомитами. Кроме того нормальный рострально-каудальный паттерн сомитной мезодермы нарушен. В результате производные сомитной мезодермы в особенности осевой скелет сильно дизорганизованы. Предполагается, что lunatic fringle модулирует передачу сигналов Notch в сегментной пластинке мезодермы и тем самым регулирует сомитогенез и одновременно формирование ростро-каудального паттерна сомитов (Evrard et al., 1998).
Conlon RA, Reaume AG, Rossant J:
Notch1 is required for the coordinate segmentation of somites. Development
1995,
121:
1533–1545.
Jiang YJ, Smithers L, Lewis J:
Vertebrate segmentation: the clock is linked to Notch signalling. Curr Biol
1998,
8:
R868–R871.
McGrew MJ, Pourquié O:
Somitogenesis: segmenting a vertebrate. Curr Opin Genet Dev
1998,
8:
487–493.
Keynes RJ, Stern CD:
Mechanisms of vertebrate segmentation. Development
1988,
103:
413–429.
Johnston SH, Rauskolb C, Wilson R, Prabhakaran B, Irvine KD, Vogt TF:
A family of mammalian Fringe genes implicated in boundary determination and the Notch pathway. Development
1997,
124:
2245–2254.
Cohen B, Bashirullah A, Dagnino L, Campbell C, Fisher WW, Leow CC, Whiting E, Ryan D, Zinyk D, Boulianne G et al.:
Fringe boundaries coincide with Notch-dependent patterning centres in mammals and alter Notch-dependent development in Drosophila. Nat Genet
1997,
16:
283–288.
de la Pompa JL, Wakeham A, Correia KM, Samper E, Brown S, Aguilera RJ, Nakano T, Honjo T, Mak TW, Rossant J, Conlon RA:
Conservation of the Notch signalling pathway in mammalian neurogenesis. Development
1997,
124:
1139–1148.
Kusumi K, Sun ES, Kerrebrock AW, Bronson RT, Chi DC, Bulotsky MS, Spencer JB, Birren BW, Frankel WN, Lander ES:
The mouse pudgy mutation disrupts Delta homologue DII3 and initiation of early somite boundaries. Nat Genet
1998,
19:
274–278.
•
Forsberg H, Crozet F, Brown NA:
Waves of mouse Lunatic fringe expression, in four-hour cycles at two-hour intervals, precede somite boundary formation. Curr Biol
1998,
8:
1027–1030.
•
McGrew MJ, Dale JK, Fraboulet S, Pourquié O:
The lunatic fringe gene is a target of the molecular clock linked to somite segmentation in avian embryos. Curr Biol
1998,
8:
979–982.
•
Aulehla A, Johnson RL:
Dynamic expression of lunatic fringe suggests a link between notch signaling and an autonomous cellular oscillator driving somite segmentation. Dev Biol
1999,
207:
49–61.
Palmeirim I, Henriques D, Ish-Horowicz D, Pourquié O:
Avian hairy gene expression identifies a molecular clock linked to vertebrate segmentation and somitogenesis. Cell
1997,
91:
639–648.
Micchelli CA, Rulifson EJ, Blair SS:
The function and regulation of cut expression on the wing margin of Drosophila: Notch, Wingless and a dominant negative role for Delta and Serrate. Development
1997,
124:
1485–1495.
Holland LZ, Kene M, Williams NA, Holland ND:
Sequence and embryonic expression of the amphioxus engrailed gene (AmphiEn): the metameric pattern of transcription resembles that of its segment polarity homolog in Drosophila. Development
1997,
124:
1723–1732.
Wehrli M, Tomlinson A:
Independent regulation of anterior/posterior and equatorial/polar polarity in the Drosophila eye; evidence for the involvement of Wnt signaling in the equatorial/polar axis. Development
1998,
125:
1421–1432.
Bettenhausen B, Hrabé de Angelis M, Simon D, Guénet J, Gossler A:
Transient and restricted expression during mouse embryogenesis of DII1, a murine gene closely related to Drosophila Delta. Development
1995,
121:
2407–2418.
Dunwoodie SL, Henrique D, Harrison SM, Beddington RS:
Mouse DII3: a novel divergent Delta gene which may complement the function of other Delta homologues during early pattern formation in the mouse embryo. Development
1997,
124:
3065–3076.
Zeilder MP, Perrimon N, Strutt DI:
Polarity determination in the Drosophila eye: a novel role for Unpaired and JAK/STAT signaling. Genes Dev
1999,
13:
1342–1353.
Bishop SA, Klein T, Martinez Arias A, Couso JP:
Composite signaling from Serrate and Delta establish leg segments in Drosophila through Notch. Development
1999,
126:
2993–3003.
Jen W-C, Gawantka V, Pollet N, Niehrs C, Kintner C:
Periodic repression of Notch pathway genes governs the segmentation of Xenopus embryos. Genes Dev
1999,
13:
1486–1499.
Takke C, Campos-Ortega JA:
her1, a zebrafish pair-rule like gene, acts downstream of notch signalling to control somite development. Development
1999,
126:
3005–3014.
del Barco Barrantes I, Elia AJ, Wünsch K, Hrabe De Angelis M, Mak TW, Rossant J, Conlon RA, Gossler A, de la Pompa JL:
Interaction between Notch signaling and Lunatic fringe during somite boundary formation in the mouse. Curr Biol
1999,
9:
470–480.
Regulation and function of Lunatic Fringe in vertebrate somitogenesis
Роль сигналов Notch в сомитогенезе впервые выявлена с помощью генного таргетинга у мышей Notch1[48] и затем продемонстрирована у кур, лягушек и рыб (reviewed in [49][50]). Сомитогнез(reviewed in [51]) (Fig. 3). Несколько генов идентифицировано в сомитах. Эти молекулярные маркеры сегментации показывают, что вновь формирующиеся сомиты уже подразделены на отдельные передение и задние типы клеток .
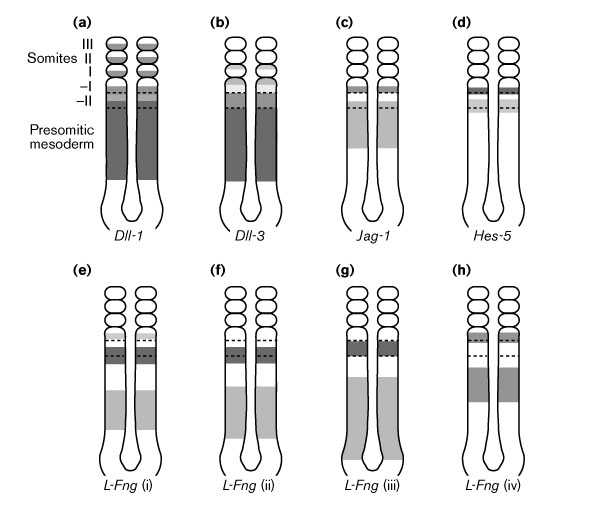
Рис. 3 Экспрессия L-Fng, Notch лигандов, и Hes-5 во время сомитогенеза у мышей. Экспрессия Notch1 и Notch2 (не показано) модулируется во время сомитогенеза. Затемнение соответствует относительным уровням экспрессии. Границы между презумптивными сомитами не обнаруживаются в пресомитной мезодерме, хотя взаимоотношения между паттернами экспрессии генов и будущими границами сомитов их выявляют. (a)Delta-like 1 (Dll-1) экспрессируется в задних частях сформированных (–I) и во вновь формируемыми (I) сомитах [63][64]. (b)Delta-like 3 (Dll-3) экспессируется в передних частях формируемых и вновь сформированных сомитов [64]. (c)Jagged/Serrate-1 (Jag-1) экспрессируется в задних частях формирующихся сомитов [21•][53]. (d)Hes-5 экспрессируется в задних частях или на задней границе формирующихся сомитов [22•][54]. (e–h)L-Fng экспрессия является динамичной [56•]. i–iv представляет различные ступени в цикле L-Fng экспресии. Имеются некоторые различия между профилями экспрессииL-Fng у мышей и кур и некоторы е различия между профилями разных групп [52][53][56•][57•][58•] но в целом экспрессия L-Fng движется волнами к переднему (ростральному) концу сомитной мезодермы и эти волны становятся уже по мере их продвижения вперед. У мышей, но не у кур наиболее передние волны экспрессииL-Fng исчезают прежде чем вновь сформированный сомит становится идентифицируемым. Figure 3 Expression of L-Fng, Notch ligands, and Hes-5 during mouse somitogenesis. The expression of Notch1 and Notch2 (not shown) is also modulated during somitogenesis. Shading approximates relative levels of expression. Borders between presumptive somites are not identifiable in the presomitic mesoderm, hence the relationships between gene expression patterns and future somite boundaries are approximate. (a)Delta-like 1 (Dll-1) is expressed in the posterior of forming (–I) and newly formed (I) somites [63][64]. (b)Delta-like 3 (Dll-3) is expressed in the anterior of forming and newly formed somites [64]. (c)Jagged/Serrate-1 (Jag-1) is expressed in the posterior of the forming somite [21•][53]. (d)Hes-5 is expressed in the posterior or at the posterior border of the forming somite [22•][54]. (e–h)L-Fng expression is dynamic. (This depiction is adapted largely from [56•].) i–iv represent different steps in a cycle of L-Fng expression. There are some differences between the reported L-Fng expression profiles in mice and chicks and there are also some differences between the profiles described by different groups [52][53][56•][57•][58•] but, in general, L-Fng expression appears to move in waves towards the anterior (rostral) end of the presomitic mesoderm, and these waves narrow as they move anteriorly. In mice (but not in chicks) the most anterior wave of L-Fng expression fades away before the newly formed somite is morphologically identifiable.
Нарушение передачи сигналов Notch не приостанавливает сомитогенез, но вызывает дизорганизацию этого процесса, сомитогенез задерживатеся, формируется меньше границ между сомитами и эти границы расположены нерегулярно (reviewed in [49][50]). Молекулярно, некоторая сегментная периодичность в паттернах экспресссии генов сохраняется, но экспрессия границ более диффузна. Наиболее существенным инициальным дефектом является разрушение A–P компартментализации в формирующихся сомитах: гены, обычно экспрессирующиеся или в передених или в задних клетках, теперь экспрессируются изменчиво по всей ширине сомита. Неясно, существует ли независмая потребность в сигналах Notch как для формирования границ, так и A–P компартментализации, или один из этих фенотипов является вторичным.
Экспрессия в виде полос Lunatic Fringe (L-Fng) в пресомитной мезодерме указывает на то. что он играет важную роль в сомитогенезе [52][53] (Fig. 3) . Мутанты L-Fng имеют дефекты в A–P подразделении и в формировании границ сомитов [21•][22•]. L-Fng мутанты также влияют на экспрессию Hes-5 [22•],который является геном-мишенью для Notch [54]. Нарушение сигнализациии[55] Notch ведет к более диффузной экспрессии Notch и Notch лигандов. Хотя эти нарушения в экспрессии генов пути Notch могут быть результатом нарушения петли обратной связи, сходной с таковой в крыльях и глазах Drosophila, однако возможно также, что они являются вторичным следствием дефектов формирования сомитов.
L-Fng экспрессия в виде полос в пресомитной мезодерме на ранних стадиях сомитогенеза скорее, чем любого другого члена пути Notch [52][53][56•][57•][58•] (Fig. 3). Более того, L-Fng регулируется молекулярными часами, которые функционируют во время сомитогенеза. Изучение гена hairy у кур показало существование этого аутосомного клеточного осцилятора, который управляет циклическими волнами экспрессии hairy в пре-сомитной мезодерме [59]. L-Fng экспрессия испытывает сходные осциляции [56•][57•][58•], и так как L-Fng, а не hairy осциляции зависят от синтеза белка, кажется наиболее вероятным, что L-Fng регулируется с помощью hairy[57•]. Выявляется критическая связь между клеточным осцилятором и регуляцией сигналов Notch. Однако, L-Fng экспрессия в пресомитной мезодерме динамична и неясно, какой аспект паттерна экспрессии наиболее важен для сомитогенеза. В формировании (–I) пре-сомита, L-Fng экспрессия примерно в половине сомита, но не в –II пресомите, L-Fng экспрессия на всю ширину сомита (Fig. 3). Хотя экспрессия в половине сомита может быть сравнена с постулируемой ролью A–P компартментализации, L-Fng экспрессируется в переденей половине –I сомита кур [22•][57•], и в задней половине –I сомита у мышей [52][56•], и, по-видмому, мало вероятно, что мыши и куры используют различные механизмы A–P компартментализации.
Conclusions
fringe модулирует передачу сигналов Notch так, что активация Notch оказывается позиционированной вдоль границы экспрессии fringe.
Схематическое представление экспрессии c-hairy1 в ПСМ в отношении индивидуальных пресомитных клеток. А. 4 диаграммы сегментной пластинки во время формирования следующего сомита показывают статические интервалы волны экспрессии с-рфшкн1, которая проходит черех ПСМ в ростро-каудально каждые 90 мин. Экспрессия начинается в широком каудальном домене, который сужается по мере его продвижения кпереди и наконец ограничивается каудальным доменом следующего сомита, который будет сформирован. Таким образом, каждая индивидуальная клетка ПСМ (точка) проходит фазу включения и выключения c-hairy1 во время каждой осцилляции. S0, формирующийся сомит, S1 только что сформированный сомит, SII,первый из сформированных сомитов. В: история пресомитной клетки (точка) в ПСМ: с момента ее инкорпорации в сомит. Этот временной интервал соответствует формированию 12 сомитов. Во время ее нахождения в ПСМ клетка подвергается 12 циклам экспрессии с-рфшкн1. Так как начало циклической экспрессии РНК инициируется в стволовых клетках полоски, то сомитные клетки передних сомитов испытывают немного клеточных уиклов прежде, чем они вступят в ПСМ, чем те, что станут каудальными сомитами. Таким образом общее количество циклов экспрессии в кдлетках определенного сомита будет отличным и будет соответствовать их расположению вдоль оси тела. Эти осцилляции следовательно предопределят связь часов с сегментацией сомитов и возможно с регионаолизацией сомитов вдоль переднезадней оси.
МОДЕЛИ ГЕНЕРАЦИИ МЕТАМЕРНОГО ПАТТЕРНА
Сегментный препаттерн в пресомитной мезодерме
Классической моделью предполагается, что клетки, располагающиеся в пресомитной мезодерме (ПСМ) уже предетерминированы (prepatterned) вдоль переднезадней оси в отношении их последующей группировки в сомиты. Эта способность генерируется наиболее каудальной часть ПСМ, в области ее перехода от первичной полоски, а позднее от хвостовой почки. Иными словами периодичность сегментации является отражением поступления прегруппированных сомитогенных клеток из полоски в ПСМ. Модель "препаттерна" однако неспособна объяснить механизм периодичности этого процесса препаттерна.
Модель часов и фронта волны
Модель постулирует, что позиционная информация, отличающаяся в каждой пресомитной клетке, является функцией вумудщзьутефд времени. В некий критический момент развития значения этих переменных таковы, что клетки подвергаются внезапным поведенческим изменениям, например, клеточной адгезивности. Так как разитиве следует вдоль гладкого переднезаднего градиента, то наблюдаемые поведенческие изменения клеток должны быть связаны с порогом позиционной информации, занчения в определенный отрезок времени будут проявляться как гладкая волна. Эта "волна", по-видимому, оперирует сочетанно с внутриклеточным осциллятором, постоянно оперирующим в пресомитных клетках. Осцилятор должен быть связан по фазе в соседних клетках и должен запускать ответ на начало поведенческих изменений, способствуя, а затем ингибируя реакцию на фрон волны. Иными словами должна быть гладкая переднезадняя онтогенетическая волна изменения поведения клеток, но осцилирующая природа реакции обусловливает поступление дискретных последовательных популяций.
Модель Мейнхарда
Она также базируется на идее осциляторного механизма, опреирующего в пресомитных клетках и продуцирующего отличающие состояния между клетками, такие как передние и задние. Предполагается, что периодичность каждой полной осциляции должна быть периодом времени, достаточным для формирования сомита, и что клетки с оппозитными "состояниями" не должны смешиваться. Согласно этой модели эти флюктуации между состояниями клеток стабилизирутся на передней границе ПСМ, когда зачаток сомита отделяется и клетки со сходными состояниями группируются отдельно.
Сегментация нуждается в формировании межсегментных границ. Для этого Мэйнхард постулирует третье состояние клеток межсомитных границ.
Молекулярные часы и сегментация
Показано, что экспрессия c-hairy1 идентифицирует молекулярные часы для сомитогенеза, которые тикают в клетках ПСМ. Клетки в передней половине формирующегося сомита не экспрессируют c-hairy1, тогда как клетки в будущей задней половине сохраняют экспрессию c-hairy1. Динамика экспрессии c-hairy1 очень напоминает осцилятор в модели Мэйнхарда, детерминирующего качественные особенности полусомитов.
Сигналы Notch лежат ниже часового механизма. В ростаральной части ПСМ кроме того экспрессируются гены Delta1 и Delta3, которые оказываются ограничены передними и задними частями сомита и участвуют в преденезадней спецификации сомита.
Ген lunatic fringle также имеет осцилирующую экспрессию в ПСМ. Этот ген участвует в пути передачи сигналов Notch. Он действует, по-видимому, ниже чем c-hairy1, но выше Notch пути, определяя места формирования границ.
Клеточный цикл как часы
Штерн с сотр. наблюдали после однократного пульсового воздействия тпловго шока аномалии сегментации повтрояющиеся трижды с интервалами в 5-7 сомитов. Периодичность этих аномалий коррелирует с продолжительностью клеточного цикла. Более того они показали существование синхронности клеточных циклов некоторых клеток в ПСМ. Эти клетки детерминированы, чтобы принадлежать одному сомиту.
У большинства позвоночных каждый уровень вдоль переднезадней оси, затылочный, шейный, торакальный, поясничный и кресцовый обычно представлен 5-7 позвонакам и, специфичными для каждой области. Следовательно, клеточные циклы, частичносинхронизированые в ПСМ, м могут контролировать переход от генерации сомитов, типичных для одной области, к генерации для следующей области.
"Einbahnstrasse" модель
Модель Duboule напрямую связывает клеточный цикл с временной регуляцией региональной идентичности с помощью Нох генов.
Предполагаемый механизм контроля связан с накоплением белков, которые специфически связываются с высоким сродством с ДНК в 5э области Нох комплекса. Это блоекирует доступ к механизму транскрипции, но при последующих деления клеток эти транскрипционные антогонисты прогрессивно оттитровываются, что ведет к транскрипции все более 5э генов в комплексе. Модель м.б. одинаково применима как к пространственному, так и временному контролю экспрессии НОх генов в ПСМ. Таким образом. последовательные деления стволовых клеток в примитивной полоске могут регулировать последовательную активацию Нох генов в клетках, которые проникают в ПСМ. Клетки ПСМ, которые возникают раньше и располагаются впереди будут экспрессировать 3' гены, тогда как все больше 5' генов будут экспрессировать ПСМ клетки, продуцируемые позднее и располагающиееся более каудально. Следовательно, будет предопределяться региональная судьба сомитных производных.
Сомитная регионализация и часы сегментации
В модели Мэнхарда число осциляций, испытываемое каждой клетокй, предопредляет сегмент-специфическую судьбу производных (шейную, торакальную и т.д). Предполагается, что механизм ритмических движений стрелок часов может участвовать в формировании сомитных единиц и региональных доменов.
Предполагается, что осциляции начинаются очень рано в развитии сомитных клеток. Региональная детерминация клеток параксиальной мезодремы вдоль оси , по-видимому, происходит непосредственно после того, как они покидают первичную полоску )или хвостовую почку) и вступают в ПСМ. Клетки ПСМ в которых осциллируют с-hairy1 и lunatic fringle, уже детерминированы в отношении их будущей переднезадней судьбы и локализации. Следовательно в моделях Вгищгду и Ьуштрфкве часы сегментации функционируют в клтках до вступления их в ПСМ, т.е. в сомитных стволовых клетках первичной полоски. Подтверждено, что часы сегментации уже активны в сомитных стволовых клетках, в ростаральной части полоски. Число осцилляций прямо соответствует их положению вдоль переднезадней оси (Рис. 4)
Кальциевые часы в ПСМ?
На рыбках данио показано, что во время гаструляции и сомитогенеза эмбрионы обладают периодической серией межклеточных дальнодщейтвующих волн кальция с частотой 5-10 в минуту. Более того эти пульсы исходят из удаленных локусов на разных стадиях развития, от узелка и области хвостовой почки во время сомитогенеза. Связаны ли они с сомитогенезом, пока неясно. Прямая связь маловероятна. Эти волны указывают на существание межклеточных сообщений очевидно через щелевые соединения, которые могут обеспечивать механизмы пространственной и временной координации высоко локальных процессов в больших клеточных доменах.
(Рис.1.) | Somitogenesis in mouse and zebrafish embryos.
(Рис.2.) | Components of the Notch signalling pathway that might be involved in somitogenesis.
(Рис.3.) | Expression pattern of segmentation genes in the presomitic mesoderm.
(Рис.4.) | Genetic evidence for the requirement of Mesp2 in establishing rostrocaudal polarity.
(Рис.5.) | Manipulation of an Fgf signal alters somite size in zebrafish.
THE MAKING OF THE SOMITE: MOLECULAR EVENTS IN VERTEBRATE SEGMENTATION Yumiko Saga & Hiroyuki Takeda Nature Reviews Genetics 2, 835 -845 (2001)
Во время сомитогенеза пресомитная мезодерма (PSM) м.б. подразделена, по крайней мере на две отдельные области: область I (задняя часть PSM) и область II (передняя PSM), которые соответствуют двум различным состояниям клеток, состоянию I и состоянию II.
Не обнаруживается признаков сегментной спецификации на молекулярном и клточном уровне в области I, однако PSM клетки обладают ростро-каудальной полярностью и становятся компетентными сегментами, когда они достигают области II.
Циклическая экспрессия генов, которая отражает лежащие в основе сегментации часы, транслируется в активность Notch, которая делает осциляции соседних PSM клеток синхронизированными в области I.
Экспрессия delta-like 1 (Dll1) в области II, которая регулируется с помощью mesoderm posterior 2 (Mesp2) на Notch сигнальном пути, детерминирует рострокаудальную полярность сомитов.
Передача сигналов Notch м. зависеть или не зависеть от presenilin 1 (Psen1); первые участвуют в индукции экспрессии Dll1, а последние в ингибировании экспрессии Dll1. Mesp2 м. стимулироватьингибирующий путь и супрессировать путь индукции.
Переход от состояния I к состоянию II контролируется с помощью уровня передачи сигналов fibroblast growth factor (Fgf). Высокий уровень активации Fgf в задней части PSM поддерживает клетки PSM в незрелом состоянии, тогда как низкие уровни Fgf ускоряют процесс созревания PSM клеток в передней части PSM.
Благодаря активации Fgf домен отступает перед наступлением сомитгенеза, причем фронт волны, который является промежуточной фазой между областью I и областью II, постепенно отходит назад. Итак, периодические взаимодействия между фронтом волны и волной осцилляций и создают регулярно расположенные границы сомитов.