Большинство цитоплазматичеких органелл, таких как эндоплазматический ретикулем, аппарат Гольджи, лизосомы, и большая часть цитозоля элимнируется из созравающих спематид во время спермиогенеза. Элиминации из каудальных частей сперматид осуществляется в основном через резидуальные (остаточные) тела. Резидуальные тела формируются из цитоплазматических долей удлинняющихся сперматид, отделяются от сперматид перед высвобождением спермиев (spermiation) и фагоцитируются клетками Сертоли. Однако цитоплазма головной части поздних сперматид отделяется формированием заднего кольца от цитоплазмы концевой части.
Вокруг головки поздней сперматиды образуются цитоплазматические выпячивания в инвагинации плазменных оболочек клеток Сертоли (тубулобульбарные комплексы, ТБК). Они описаны у многих млекопитающих , включая и человека. Они состоят из двух элементов: тубулярной структуры в 2-4 µm в длину и 50 &nano;m в диаметре и балонообразной терминальной бульбарной структуры 1 µm в диаметре. Около бульбарной части располагаются лизосомы и фагоцитарные узелки клеток Сертоли.
Во время ступеней 18-19 спермиогенеза электроно-прозрачная область дорсального кортекса передней акросомы постепенно уменьшается и наконец акросома конденсируется и становится уменьшенной в своем размере. Одновременно с этим формируются тубобульбарные комплексы (ТБК) из головной порции поздних сперматид, выпирающие в окружающие клетки Сертоли.
Акросомальный гликопротеин MN7, располагавшийся в задней части акросом во время стадий 8-18, внезапно перемещается в переднюю чать акросомы на стадии 19. Сходным образом и другие акросомальные белки, такие как SP-10 и АМ50, также перераспределяются в акросоме.
Затем как PAS-реактивность, так и mMN7-иммунореактивность обнаруживаются как в ТБК, так и в акросоме. Следовательно, акросомальное содержимое включается в ТБК. (Tanii et al 1999) На некоторых срезах видно, что акросома проникает в ТБК-подобную структуру обычно же ТБК содержат уплотненный материал, сходный с акросомальным. В этом случае вокруг тубул обнаруживаются микрофиламенты и покрытые щетинками углубления клеток Сертоли на дистальном конце (Рис. 4а).
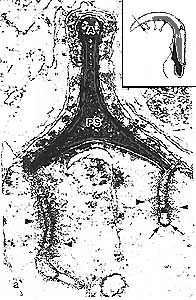
Рис. 1. Ультраструктура тубулобульбарных комплексов (ТБК) в сперматидах крыс на стадии 19 спермиогенеза. Поперечный разрез верхушки головки сперматиды. Плоскость разреза показана на вставке в верхнем правом углу. ТБК выходят из боковых краев трехугольногой головки сперматиды на стыке трех клеток Сертоли и прникают в нижнюю из них. Филаментозны материал (головки стрелок) клетки Сертоли окружает тубулярную часть ТБК. Покрытая щетинками ямка (стрелки) присутствует на дистальном конце ТБК. PS - перинуклеарное пространство, А - акросома.
Имеется, по-видимому, два типа ТБК: один связан с инкорпорацией акросомного содержимого, другой тип не связан с акросомой.
Предполагается, что акросомальные мембраны выпячиваются в некоторые ТБК на ст.18-19. Часть акросомального содержимого вытекает в ТБК , затем отшнуровывается и фагоцитируется в соседних клетках Сертоли.
Следовательно, ТБК устраняют избыток акросомального содержимого во время их конденсации. ТБК могут играть важную роль в финальном морфогенезе головок сперматид, эиминирууя избыток компоенетов, включая акросомальное содержимое и и перинуклеарную цитоплазму. Имеются и другие факторы, влияющие на морфогенез головок: конденсация ядра, микротубулярная манжетка и эктоплазматические специализации. Последние два фактора связаны с внешними физическими силами. Взаимосвязть ТБК с исчезновением электронно-прозрачной зоны в дорсальном кортексе акросом неясна. Недавно идентифицированы в сперматидах водные канальцы, аквопорины, выкачивающие воду или некоторые другие мелкие молекулы. Они могут участвовать в удалении материала из электронно-прозрачной области акросом.