Genetics and mechanics combine to guide the embryonic gut | |
|---|---|
|
Во время эмбрионального развития корректное создание 3D тела нуждается в обширном ремоделировании тканей. Слои клеток укладываются в складки и меняют свою геометрию, подвергаясь изменениям эквивалентным складыванию листа бумаги в оригами. У эмбрионов клетки, которые формируют мышечную ткань (наз. мезодермой) и ткань кишечника (энтодерму) перемещаются внутрь, а клетки наружного слоя формируют кожу. Bailles et al.1 опсали ранее неизвестный аспект того, как клетки интернализуются у Drosophila melanogaster.
Исследователи интернализации мезодермы у D. melanogaster установили молекулярную связь между клеточными особенностями (identity) и физическими изменениями, которым подвергаются клетки во время развития. Белок Twist регулирует экспрессию генов, чтобы обеспечить качественными характеристиками мышечные клетки, а клетки, которые экспрессируют Twist, сокращают свои наружные (апикальные) поверхности, сохраняя при этом контакт с соседними клетками2. Ткань прогибается в результате такого апикального сокращений, вызывая интернализацию клеток. Интернализация поэтому жестко запрограммирована в мезодерме, но она также генерирует силы, которые затрагивают соседние не мезодермальные ткани3. Насколько интернализация мезодермы сравнима с др. примерами интернализации клеток во время развития, особенно когда многие события морфогенеза происходят одновременно?
Энтодермальные клетки интернализуются в виде энтодермальной ткани вцелом - циркулярные участки приблизительно в 15 рядов клеток- мигрируют в направлении региона головы целого эмбриона. Используя микроскопию клеток вживую и экспериментальные методы, подкрепляемые математическим моделированием, авт. установи ли, что существует два самостоятельных региона энтодермы, которые отличаются механизмом своей интернализации.
Часть энтодермы, которую авт. назвали примордиальным регионом, интернализуется первой. Подобно интернализации мезодермы2, это происходит посредством процессов, которые непосредственно регулируются экспрессию генов. Экспрессия и активность белка Fog приводит к увеличению белков не мышечного myosin II (MyoII) и Rho1 в апикальном регионе клеток, приводя к апикальному сужению за счет ремоделирования клеточного цитоскелета (a филамент-подобного внутреннего каркаса в цитоплазме)4. Bailles с колл. наблюдали, что эта локальная активность Fog приводит к одновременному сужению и интернализации всех клеток примордиального региона (Fig. 1).
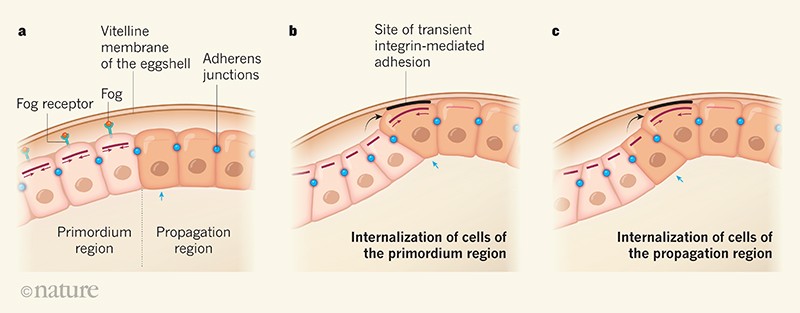 Figure 1 | The internalization of endodermal cells. a, During embryonic development of the fruit fly Drosophila melanogaster, gut-forming endodermal cells internalize, moving away from the inner layer of the eggshell (the vitelline membrane). Bailles et al.1 reveal how two endodermal regions, termed the primordium region (pink cells) and the propagation region (brown cells), internalize. Only a few cells of these regions are shown, in a cross-sectional view through the middle of the tissue, where each cell visible represents one row of cells that is perpendicular to this field of view. A cell of the propagation region is indicated by a blue arrow to aid tracking of its internalization across the panels. Cells of the primordium region secrete the protein Fog, which binds to its receptor, triggering the accumulation and activation of the protein non-muscle myosin II (pink lines, with darker shades corresponding to greater accumulation and activation). This helps to generate contractile forces (pink arrows). Structures called adherens junctions enable mechanical coupling and the transmission of force between cells. b, The cells of the primordium region simultaneously contract their outer (apical) surfaces and internalize. The primordium region transmits mechanical stress (black arrow) to cells in nearby rows of the propagation region. The cells of the adjacent row of the progagation region, only one cell of which is shown, then extend upwards towards the eggshell and transiently adhere there in a process mediated by integrin proteins. The cells of this row accumulate non-muscle myosin II. c, The first part of the propagation region internalizes after contraction of its apical surface. The entire process is repeated sequentially across this region.
Figure 1 | The internalization of endodermal cells. a, During embryonic development of the fruit fly Drosophila melanogaster, gut-forming endodermal cells internalize, moving away from the inner layer of the eggshell (the vitelline membrane). Bailles et al.1 reveal how two endodermal regions, termed the primordium region (pink cells) and the propagation region (brown cells), internalize. Only a few cells of these regions are shown, in a cross-sectional view through the middle of the tissue, where each cell visible represents one row of cells that is perpendicular to this field of view. A cell of the propagation region is indicated by a blue arrow to aid tracking of its internalization across the panels. Cells of the primordium region secrete the protein Fog, which binds to its receptor, triggering the accumulation and activation of the protein non-muscle myosin II (pink lines, with darker shades corresponding to greater accumulation and activation). This helps to generate contractile forces (pink arrows). Structures called adherens junctions enable mechanical coupling and the transmission of force between cells. b, The cells of the primordium region simultaneously contract their outer (apical) surfaces and internalize. The primordium region transmits mechanical stress (black arrow) to cells in nearby rows of the propagation region. The cells of the adjacent row of the progagation region, only one cell of which is shown, then extend upwards towards the eggshell and transiently adhere there in a process mediated by integrin proteins. The cells of this row accumulate non-muscle myosin II. c, The first part of the propagation region internalizes after contraction of its apical surface. The entire process is repeated sequentially across this region. В др. части энтодермы, которую авт. назвали регионом размножения-распространения (propagation), интернализация происходит прогрессивно, один ряд клеток за один раз. Bailles с колл. сделали неожиданное открытие, что если транскрипция супрессирована или локальный источник белка Fog потерян, то интернализация региона propagation всё ещё происходит, даже когда primordium регион уже начал сокращаться. Моделирование скорости интернализации региона propagation сравниваемое с максимумом, подсчитанной скорости диффузии Fog позволило авт. исключить диффузию Fog в качестве объяснения того, как этот процесс интернализации контролируется.
Bailles с колл. затем исследовали, могут ли механические влияния играть роль. С этой целью они физически препятствовали движению ткани, используя генетические подходы, чтобы изменить геометрию эмбриона, или использовали лекарства, подавляющие MyoII. Их эксперименты выявили, что интернализация региона propagation осуществляется с помощью механической системы позитивных обратных связей. Первоначальные, легкие клеточные деформации в результате внешних физических стрессов передаются клеткам с помощью соседей, которые подвергаются интернализации, запускается накопление MyoII в апикальной области не интернализованных клеток. Такое накопление управляет изменениями формы клеток, это приводит к дальнейшему увеличению физического стресса и дальнейшему накоплению MyoII до тех пор, пока не будет достигнуто клеточное сокращение, уровень которого вызывает интернализацию клеток. Механические связи между соседними клетками, обеспечиваемые белковыми комплексами, наз. слипчивыми соединениями, которые соединяют клетки, гарантируют, что силы будут передаваться по ткани, управляя прогрессом интернализации в регионе propagation.
Механическая регуляция развития была описана на др. экспериментальных системах5. Однако, часто трудно убедительно доказать, действуют ли механические силы на клетки, является причиной или следствием процесса развития. Хотя это заставляет оценивать механические воздействия in vivo, работа Bailles с колл. в контексте целого эмбриона увеличивает количество доказательств в пользу механических сил, как прямого регулятора развития.
Клетки в обоих энтодермальных регионах приобретают молекулярные характеристики качественных особенностей энтодермы перед интернализацией. Несмотря на это сходство, почему же эти регионы используют разные механизмы интернализации? Может быть из-за того, что если вся энтодерма будет сокращается одновременно, то геометрия эмбриона будет нарушена. Др. возможность заключается в том, что различия в механической чувствительности между этими регионами создают буфер для копировния внешних сил, которые возникают от разных, конкурентных событий развития. 
A gut feeling for cellular fate
Ирония, что идентификация механической регуляции, оказывающей критическое влияние на интернализацию региона propagation, заставляет исследовать локальную экспрессию генов. Насколько отличаются генетически primordium и propagation регионы? Напр., если бы уровни Fog были бы ниже, чем в норме, то смогло бы это открыть способность клеток в primordium регионе интернализоваться с помощью механизма, ассоциированного с propagation регионом?
Необходимо рассмотреть, как яйцо, в котором находится эмбрион, предопределяет физический контекст морфогенеза. В самом деле, оболочка яйца, как было установлено, играет большую роль в событиях интернализации тканей во время развития жука Tribolium castaneum6. Исходя из этой работы, Bailles с коллегами исследовали, может ли и у плодовой мушки оболочка яйца (слой, наз. вителлиновой мембраной) влиять на интернализацию региона propagation. Они наблюдали, что взаимодействия между эмбрионом и вителлиновой мембраной составляют источник механических сил. Клетка на острие интернализации сначала расширяется вверх в направлении оболочки яйца, затем движется резко вниз в направлении внутрь подобно волне на трибуне стадиона и этот паттерн движений повторяется по одному ряду клеток каждый раз, благодаря переходу на соседние клетки региона propagation (Fig. 1). Во время движения вверх каждая клетка временно прилипает к оболочке яйца посредством белка, наз. integrin, который экспрессируется клетками энтодермы. Такое взаимодействие апикальной поверхности энтодермальных клеток, чтобы расшириться, которое, по-видимому, вызывает небольшое количество противодействий интернализации, возможно генерирует силы, которые обеспечивают в дальнейшем позитивные ответные реакции, чтобы поднимать уровни MyoII и тем самым повышать эффективность клеточных сокращений и интернализации.
Динамика адгезии играет критическую роль в разных примерах морфогенеза7,8, а специфический механизм integrin обеспечиваемой адгезии с оболочкой яйца согласно Bailles et al. и др.6, участвует во многих онтогенетических контекстах. У T. castaneum, это мезодерма, которая экспрессирует интегрины, и их функция не усиливает механические стрессы, а скорее ограничивает смещения в ткани, когда целый эмбрион подвергается прогрессивной интернализации по отношению к не эмбриональным наружным защитным тканям6.
Будущие исследования повысят наше понимание морфогенеза на молекулярном и клеточном уровне, это поможет понять, как обычно осуществляются взаимодействия между механической и генетической регуляцией. to reveal how commonly interplay occurs between mechanical and genetic regulation.
|