Functional involvement of VAMP/synaptobrevin-2 in cAMP-stimulated aquaporin 2 translocation in renal collecting duct cellsJournal of Cell Science. 2002. V. 115. P. 3667-3674. | |
|
Функциональное участие VAMP/синаптобревина-2 в цАМФ-стимулируемом переносе аквапорина 2 в клетках собирающего протока почек Исследовали участие растворимых протеинов N-этилмалеимид-чувствительного белкового рецептора (SNARE) в цАМФ-индуцируемом экзоцитозе везикул, содержащих аквопорин 2 (AQP2), в AQP2-трансфицированных CD8-клетках почек. Методами RT-PCR и вестерн-блоттинга было подтверждено присутствие SNARE-гомологов VAMP/синаптобревина-2, синтаксина-1, синтаксина-4 и SNAP-23 в клетках CD8. Использование нейротоксина столбняка (TeNT) было эффективно в разложении синаптобревин-подобного белка, как in vitro, так и в интактных CD8-клетках, инкубированных с токсином. Воздействие TeNT на инкубированные CD8-клетки совершенно прекращало цАМФ-стимулируемый захват аквапорина 2 плазматической мембраной. При количественном анализе было выявлено, что иммунореактивность поверхности клетки к анти-AQP2-антителу возрастает по сравнению с репродукцией пептидов внеклеточной C-петли аквапорина 2. Эти результаты представляют собой первое свидетельство функционального участия VAMP-2 в цАМФ-индуцированном экзоцитозе аквапорина 2 в клетках почек.
|
Реабсорбция воды в собирающем протоке почки регулируется аквопорином 2 (AQP2) и сопряжена с процессами экзо-/эндоцитоза. Аквапорин 2 содержится во внутриклеточных везикулах, которые соединяются с апикальной мембраной основных клеток собирающего протока в присутствии антидиуретического гормона - вазопрессина (Knepper & Inoue, 1997; Klussmann et al., 2000). Главные кандидаты на роль белков, обеспечивающих специфичность поляризованной сортировки - растворимые адаптерные молекулы - протеины N-этилмалеимид-чувствительных белковых рецепторов (SNARE). Специфичность этого процесса достигается путём попарного объединения SNARE-белков в везикулах носителя - v-SNARE (Q-SNARE - Fasshauer et al., 1998) с родственными им SNARE-белками в мембране клеток-мишеней - R-SNARE (Aroeti et al., 1998; Rothman, 1994; Rothman & Wieland, 1996). Белки Q-SNARE и R-SNARE были идентифицированы в процессах захвата на всём протяжении от аппарата Гольджи до плазматической мембраны в нейронах и у дрожжей (Rothman, 1994). Вероятно, что событие, которое регулирует перенос аквапорина 2 в клетках почек, использует для экзоцитоза механизм, подобный описанному для клеток нейронов. Этой гипотезе не противоречат предыдущие исследования, которые подтвердили экспрессию SNARE-белков в клетках собирающего протока почки (Jo et al., 1995; Liebenhoff & Rosenthal, 1995; Nielsen et al., 1995; Mandon et al., 1996; Inoue et al.,1998). Однако до настоящего времени не было представлено прямых доказательств функционального участия SNARE-белков в регулируемом экзоцитозе аквапорина 2 водных каналов.
Для того чтобы исследовать, участвуют ли SNARE-белки в этом процессе, нами были использованы AQP2-трансфицированные CD8-клетки (Valenti et al., 1996), которые являются в данном случае идеальной моделью для изучения молекулярных механизмов экзоцитоза (Valenti et al., 1998; Valenti et al., 2000; Tamma et al., 2001). Нами была установлена экспрессия SNARE-белков. Кроме того, мы исследовали эффект нейротоксина столбняка (TeNT) на цАМФ-индуцированное перемещение аквапорина 2 в интактных CD8-клетках. Настоящее исследование ясно доказывает, что SNARE-белок VAMP/синаптобревин-2 (VAMP-2) функционально вовлечён в цАМФ-стимулируемый перенос аквопорина 2 в клетках собирающего протока почек. ОБСУЖДЕНИЕ
Коэффициент водной реабсорбции в собирающем протоке почки регулируется вазопрессином. Этот гормон управляет межклеточным переносом воды путём перераспределения AQP2-несущих везикул к апикальной мембране основных клеток собирающего протока.(Knepper & Inoue, 1997; Klussmann et al., 2000). Чтобы исследовать функциональную причастность SNARE-белков к процессу регулируемого перемещения инсерции водного канала, мы использовали клетки собирающего протока линии CD8 стабильно трансфицированные чувствительным к вазопрессину аквапорином 2. 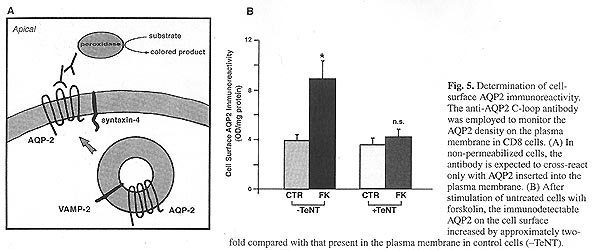 В данном исследовании, представлено доказательство, прямо подтверждающее гипотезу, что v-SNARE-белок VAMP-2 вовлечён в захват апикальной плазматической мембраной аквапорина 2 из везикул. В интактных CD8-клетках форсколин-индуцированное перемещение аквапорина 2 к апикальной мембране было прекращено при воздействии TeNT (рис. 5). Этот токсин протеолитически разлагает VAMP-2 (Link et al., 1992). Было предположено, что VAMP-2 играет ведущую роль в механизме траффикинга везикул (Sollner et al., 1993). Это доказательство основано, главным образом, на способности TeNT и ботулинических нейротоксинов разлагать VAMP-2 в различных точках (Schiavo et al., 1995), что приводит к ингибированию нейромедиатора в нейронных и нейроэндокринных клетках (Hunt et al., 1994). Регуляторная роль изоформ синаптобревина также была доказана для экзоцитозе GLUT4 (в адипосайтах), гистамина (в энтерохромаффин-подобных клетках), акросом сперматозоидов и для траффикинга H+-АТФазы (в мозговом слое собирающего протока): (Banerjee et al., 2001; Banerjee et al., 1999; Foster & Klip, 2000; Hohne-Zell et al., 1997; Martin et al., 199S; Schuiz et al., 1997). Таким образом, SNARE-белки, вероятно, выполняют универсальную роль в везикулярно-мембранном слиянии как в нейронах, так и в клетках других тканей.
В клетках почек путём количественной двойной иммунометки, была показана, колокализация VAMP-2 с везикулами, содержащими аквапорин 2 (Nielsen et al., 1995), подтверждающая роль VAMP-2 в вазопрессин-регулируемом траффикинге везикул. В том же исследовании авторы продемонстрировали, что TeNT вызывал полное разложение VAMP-2 в мембранной фракции, обогащённой внутриклеточными везикулами из почечного мозгового вещества (Nielsen et al., 1995). Jo и соавторы (1995) сообщили, что очищенные AQP2-содержащие эндосомы внутреннего мозгового слоя из почки крысы, содержат VAMP-2. Авторы продемонстрировали слияние эндосом in vitro, посредством ATФ-зависимого процесса, который значительно ингибируется, если эндосомы были преинкубированы с anti-VAMP-2-антителом или TeNT.
Несмотря на факт, что эти исследования определённо указывают на функциональную причастность VAMP-2 к AQP2-регулируемому перераспределению, прямого доказательства этой роли в интактных клетках почек пока нет. Мы попытались продемонстрировать роль VAMP-2 в поляризованной сортировке путём определения влияния TeNT клостридий на цАМФ-индуцируемый перенос в интактных CD8-клетках почек. Центральным моментом в исследовании была демонстрация того, что TeNT может проникать в клетки и разлагать эндогенно экспрессированный VAMP-2. TeNT в интактных CD8-клетках полностью прекращал цАМФ-стимулируемый захват аквапорина 2 плазматической мембраной. При количественном анализе было выявлено, что иммунореактивность поверхности клетки к анти-AQP2-антителу возросла по сравнению с репродукцией пептидов внеклеточной C-петли аквапорина 2.
Эти результаты представляют собой прямое доказательство функциональной роли VAMP-2 в цАМФ-индуцированном перераспределении аквапорина 2 в клетках почек. В отличие от влияния токсина на цАМФ-индуцированное перемещение аквапорина 2 к апикальной мембране, количество поверхностного аквапорина 2 в покоящихся клетках не понижается при выделении токсина (рис. 5). Это наблюдение позволяет предположить, что конститутивная доставка аквапорина 2 может быть не зависимой от VAMP-2.
В нейронах, VAMP-2 формирует комплекс с двумя плазма-мембранносвязанными SNARE-белками: синтаксином и SNAP-25. Они связываются вместе параллельным образом (Hanson et al., 1997), формируя связку с четырьмя завитками, два из которых "оккупированны" SNAP-25 и ещё два - соответственно VAMP-2 и синтаксином (Sutton et al., 1998). Формирование комплекса SNARE-белками само по себе является достаточным, чтобы приблизить две взаимодействующие мембраны на расстояние, делающее слияние возможным (Weber et al., 1998).
Среди известных изоформ синтаксина, только синтаксин-1 и синтаксин-4 связываются с VAMP-2 с высокой аффинностью (Calakos et al., 1994; Pevsner et al., 1994). Синтаксин-4 локализован в апикальной мембране основных клеток собирающего протока (Mandon et al., 1996), из чего можно предположить, что он представляет собой аналоговый белок, взаимодействующий с VAMP-2, который был найден в везикулах, содержащих аквапорин 2 (Jo et al., 1995; Nielsen et al., 1995). Наличие VAMP-2, синтаксина-1A и синтаксина-4 исследовалось методами RT-PCR и вестерн-блоттинга в CD8-клетках. В большом количестве VAMP-2 был найден во внутриклеточных везикулах, в то время, как и синтаксин-1, и синтаксин-4 были найдены в больших количествах во фракции плазматической мембраны. Дальнейшие исследования будут направлены на изучение того, взаимодействует ли VAMP v-SNARE-2 с синтаксином-1 или синтаксином-4 для опосредованного захвата аквапорин 2 .
SNAP-25 участвует в слиянии на позднем этапе (Banerjee et al., 1996; Mehta et al., 1996; Rossi et al., 1997). В клетках внутреннего мозгового слоя собирающего протока обработка ботулиническим токсином E, разлагающим SNAP-23 у крыс, понижала количество H+-АТФазы, перемещённое к апикальной мембране, примерно до 52%, демонстрируя, что SNAP-23 играет критическую роль в регуляции H+-АТФазы экзоцитоза (Banerjee et al., 2001). Мы полагаем, что SNAP-23, обнаруженный в больших количествах во фракции плазматической мембраны клеток CD8, действует аналогично в клетках почек. Дальнейшее исследование необходимо, чтобы оценить роль специфических SNARE-белков в динамике формирования комплекса.
Подводя итог, можно сказать, что результаты данного исследования, во-первых, явно доказывают непосредственное участие VAMP v-SNARE-2 в стыковке и слиянии везикул, содержащих аквапорин 2, с плазматической мембраной интактных AQP2-трансфицированных CD8-клеток; во-вторых, создают основу для понимания механизма и регуляции переноса аквапорина 2 в ответ на цАМФ-агент
|