Mara C. Duncan and Gregory S. Payne TRENDS in Cell Biology Vol.13 No.5 May 2003
Clathrin-coated vesicles (CCVs) play important roles in
nutrient uptake, downregulation of signaling receptors,
pathogen invasion and biogenesis of endosomes and
lysosomes. Although detailed models for endocytic
CCV formation have emerged, the process of CCV for-mation
at the Golgi and endosomes has been less
clear. Key to endocytic CCV formation are proteins
containing related phosphoinositide-binding ENTH and
ANTH domains. Now, recent studies have identified
novel ENTH/ANTH proteins that participate in
CCV-mediated traffic between the trans-Golgi Network
(TGN) and endosomes and have defined a molecular
basis for interaction with AP-1 and GGA adaptors in cla-thrin
coats of the TGN/endosomes. Thus, ENTH/ANTH
domain proteins appear to be universal elements in
nucleation of clathrin coats.
(Табл .1.) | Examples of clathrin adaptors and their interactions.
Сноски: Cited reviews [1-3] contain original references for the indicated interactions b
Yeast and mammalian homologs of adaptors and accessory proteins are positioned in the same row c
For interaction motifs, F is a large hydrophobic amino acid d
For interaction motifs, X is any amino acid e
LL is dileucine-type sorting signal f
T.D. is the clathrin N-terminal domain g
C.B. are clathrin-binding motifs corresponding to the general consensus pL(/)p(/), where p is a polar amino acid and (/) is a hydrophobic amino acid [30] h
AcF is the acidic-phenylalanine motif described in Fig. 3 i
AcLL is acidic dileucine sorting signal
ENTH/ANTH proteins and clathrin-mediated membrane budding Valerie Legendre-Guillemin, Sylwia Wasiak, Natasha K. Hussain, Annie Angers, and Peter S. McPherson JCS 2004 117: 9-18.
Clathrin-coated vesicles (CCVs) интернализуют эндоцитозированный материал плазматической мембраны и транспортируют также груз из trans Golgi network (TGN) в endosome/lysosome систему. Недавно выявлены детали многочисленных белков, участвующих в отпочковывании клатрин-опосредованных пузырьков. В работе на , Peter McPherson и др. обсуждают роль двух родственных модулей, присутствующих у некоторых из этих белков: ENTH домен сначала был идентифицирован в связанном с клатрином белке epsin; a ANTH домен характеризовал др. связанный с клатрином белок, AP180. Оба домена связывают мембранные фосфоинозитиды, позволяя др. областям белков участвовать в сборке клатрина. Механизмы связывания различны, однако домен ENTH, но не ANTH, по-видимому, способен вызывать искривление мембраны. ENTH/ANTH белки м. выполнять и дополнительные функции: увеличиваютя доказательства, показывающие, что они действуют как груз-специфические адапторы. Более того, домены соединяются с микротрубочками и м. т. о. прикреплять клатрином покрытые мембраны к цитоскелету. Недавно было показано, что ENTH/ANTH белки м. соединяться с адапторными белками, такими как AP-1 и GGA, это особенно важно, т.к. подтверждает, что домены это фактически универсальные компоненты кухни (machinery) отпочковывания клатрин-опосредованных пузырьков.
Clathrin-coated vesicles (CCVs) являются эволюционно законсервированными переносчиками белков, которые обеспечивают эндоцитоз и перенос из TGN и эндосом в клетках эцукариот. Образование CCV обеспечивается сочетанной рекрутацией белков, которые формируют оболочечные структуры на соотв. мембранах, набором грузовых белков в coated мембраны, инвагинацией мембраны и обязательным отделением полностью сформированных покрытых оболочкой пузырьков. Достигнут существенный погресс в деле понимания молекулярных основ каждой ступени процесса эндоцитоза. Три класса белков участвуют в формировании CCV: clathrin, который образует многогранный (polyhedral) наружный покров оболочки, но не обладает врожденным сродством к мембранам; адапторы, это белки, которые соединяют клатрин с мембраной путем присоединения к клатрину phosphoinositides и/или трансмембранных грузовых (cargo) белков; и дополнительные белки, обозначаемые как акцессорные факторы. На TGN и эндосомах, роль клатрина и адапторов кажется сходной с из ролью в эндоцитозе. Однако, значительно меньше известно об адапторных и акцессорных белках, большинство их отлично от эндоцитотических adaptors/accessories, и их поставка в мембраны механистически отлична от поставки эндоцитотических оболочек. Более того, довольно мало известно о сети взаимодействий между покровными белками, грузом и мембраной, ответственной за образование TGN/endosome CCV.
Идентифицированы новые кандидаты на роль клатриновых адапторов на TGN/эндосомах. Их физические характеристики и партнеры по взаимодействию обнаруживают строгие параллели с адапторами, участвующими в эндоцитозе. Здесь основное внимание уделяется поставке оболочек на TGN и эндосомы.
Nucleation of endocytic clathrin coats
Эндоцитотические адапторы м.б. подразделены на два больших класса; мономерные адапторы, такие как AP-180, epsin1 и β-arrestins, и гетеротетрамерный адапторный комплекс AP-2 (Табл. 1). Мономерные адапторы взаимодействуют с клатрином, мембранными phosphoinositides и в некоторых случаях с грузом. AP-2 обладает всеми тремя из этих активностей. Оба адапторных м. соединяться с акцессорными факторами и др. адапторами. В AP-2, accessory/adaptor белки взаимодействуют с α и β2 субъединицами C-терминальных ‘ear’ доменов посредством коротких пептидных мотивов (Табл. 1). Определение этих связывающих мотивов предоставляет мощное средство для идентификации новых компонентов эндоцитотической кухни (machinery). Хотя большинство адапторов в отдельности м. способствовать сборке клатрина в покров-подобные решетки (cages) in vitro, однако, недавние находки говорят о том, что in vivo имеется серия кооперативных взаимодействий между мембраной, адапторами и клатрином, которые лежат в основе образования оболочки (Рис. 1a). Напр., с липидными монослоями в качестве матриц для рекрутирования клатрина, AP180, но не AP-2, стимулирует сборку клатрина, зависимую от phosphatidylinositol (4,5)-bisphosphate [PI(4,5)P2]. Однако, и AP180 и AP-2, которые взаимодействуют др. с др., и необходимы для генерации характеристик искривленности формирующихся CCVs.
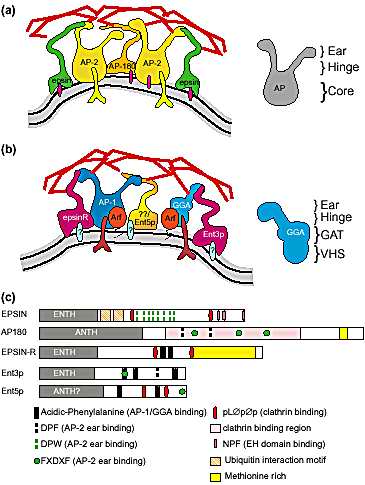
Fig. 1. Schematic views of CCV formation and ENTH/ANTH domain proteins (a)
CCVs at the plasma membrane. Epsin1, AP180 and AP-2 interact with each other,
clathrin (in red) and phosphatidylinositol (4,5)-bisphosphate [PI(4,5)P2 ] (purple
ovals). Also shown are receptors (green inverted Ys) interacting with AP-2 and
the plasma membrane (thick gray line) (b) CCVs at the TGN/endosomes. New
mammalian and yeast ENTH/ANTH domain proteins interact with adaptors. Blue
ovals represent phosphoinositides, likely to be PI(4)P for epsinR and unidentified
for Ent3p and Ent5p. Question marks in Ent5p indicate the possibility of a
mammalian homolog. Also shown are ARF GTPases that are necessary for AP-1,
GGA and epsinR membrane association [3,19]. Although AP-1 and GGAs are
shown in the same clathrin structure, in vivo they could be in distinct complexes.
(c) Domain organization of ENTH/ANTH domain proteins. C-terminal sequences
that match the consensus for AP-2, clathrin and EH domain binding are indicated.
EH domains are present in a number of endocytic proteins and recognize the pep-tide
motif NPF in proteins such as epsin1 [24]. Ubiquitin-interacting motifs (UIMs)
potentially mediate interaction with ubiquitinated cargo or endocytic machinery
[25]. With the exception of the epsinR clathrin-binding motifs, the functionality of
the clathrin- and AP-2-binding sites in epsinR, Ent3p and Ent5p are uninvestigated.
The function of the methionine-rich sequences in epsinR and AP180 are unknown.
Др. важный фактор искривленности мембраны это epsin1. Epsin1 напоминает AP180 по своей молекулярной архитектуре (Рис. 1c). Оба белка имеют структурно родственные N-терминальные PI(4,5)P2-связывающие домены, первоначально обозначенные как epsin N-terminal homology (ENTH) домены, сопровождаемые довольно неструктуированными С-терминальныи сегментами, несущими clathrin- и AP-2 ear-связывающие мотивы. Т.к. сходно упакованные ENTH домены epsin1 и AP180 соединяются с PI(4,5)P2 в разных сайтах и возможно выполняют разные функции, поэтому McMahon и др. предлагают подразделить два домена как ENTH и ANTH. Домен ANTH у CALM, не-нейронального гомолога AP180, соединяется с PI(4,5)P2 на его поверхности через законсервированные богатые лизином последовательности (Рис. 2a). Epsin1 лишен этого богатого лизином мотива и соединяется с inositol (1,4,5)-trisphosphate (IP3 ) в глубокой basic борозде, создаваемой при формировании дополнительной amphipathic спирали (helix 0) на N-конце домена ENTH (Рис. 2b).
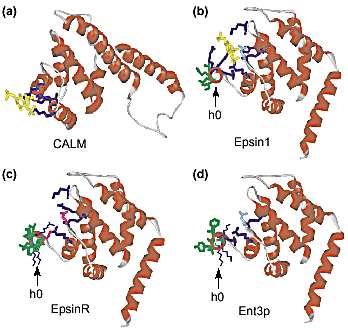
Fig. 2. ANTH and ENTH domain structure. (a) The CALM ANTH domain bound to
inositol (1,4,5)-trisphosphate (IP3) (yellow) [7]. For simplicity, only the first eight
helices of the ten-helix structure are shown. Basic amino acids contacting IP3
are diagrammed in dark blue and fall within the proposed ANTH consensus
(K/G)A(T/I)X6 (P/L/V)KXK(H/Y) [9]. (b) The epsin1 ENTH domain bound to IP3 [9].
Orientation is the same as in (a) showing the overall structural similarity of ENTH
and ANTH domains and the different modes of phosphoinositide binding. Instead
of an ANTH domain consensus, espin1 contains a proposed ENTH domain consen-sus
(D/E)ATX(D/E)PWGP in the same region of the domain [9]. The view down the
axis of helix 0 emphasizes the amphipathic arrangement of amino acids in this
helix. Hydrophobic amino acids in helix 0 are green, basic amino acids contacting
IP3 are dark blue, and a conserved asparagine is light blue. (c) ENTH domain of
epsinR modeled on espin1 ENTH. Two groups modeled the epsinR ENTH domain
based on the ligand-bound epsin1 ENTH structure and predicted an amphipathic
helix 0 [17,18]. However, epsinR exhibits substitutions in key residues in epsin1
involved in coordinating IP3; in particular epsinR contains aspartates in place of a
key arginine (R8) and asparagine (N30) in the epsin1-binding pocket. Basic amino
acids conserved with epsin1 IP3-binding residues are diagrammed with thick dark
blue side chains. Acidic amino acids replacing epsin1 IP3-binding residues have
thick red side chains. The presence of these acidic amino acids reduces the overall
positive potential of the binding site [18]. Other basic residues in helix 0 that are
not predicted to contact IP3 based on the model are shown with thin blue side
chains. (d) ENTH domain of Ent3p modeled on espin1 ENTH. Coloring scheme is
as described above. Unlike epsinR, there are no acidic amino acids replacing
potential IP3-binding residues and the overall disposition of amino acids con-served
with epsin1 IP3-binding residues is different between Ent3p and epsinR.
target pool of PI(4)P for the epsinR ENTH domain might be
generated by Golgi-localized PI(4) kinase b [21], thereby
contributing phosphoinositide specificity for CCV formation
at the TGN.
Интересно, что epsin1 м. деформировать липидные мембраны, tubulating липосомы или искривленные липидные монослои in vitro, преимущественно за счет пенетрации гидрофобной стороны (face) спирали 0 в мембрану. Исходя из этих данных, предложена привлекательная модель, согласно которой нуклеация оболочки AP180 и epsin1 осуществляется за счет связывания с PI(4,5)P2-обогащенными доменами плазматической мембраны и рекрутирования AP-2 и clathrin. Вместе, AP180 и AP-2 м. в приципе управлять полимеризацией клатриновой оболочки в полиэдральную решетку, тогда как epsin1
будет облегчать сопровождаемую инвагинацию мембраны. В согласии с разными ролями AP180 и epsin1 находится и то, что только AP180 является структурным компонентом CCVs.
TGN/endosome clathrin coat formation
Два типа адапторов, ассоциированны с образованием CCV на TGN и эндосомах, являются AP-1 комплексом (с субъединицами, гомологичными таковым в AP-2) и мономерными
GGA белками (Табл. 1). AP-1 и GGAs участвуют в транспорте из TGN в эндосомы, тогда как
AP-1 сам по себе м. обеспечить перенос эндосом на TGN. В противоположность AP-2, AP-1 и GGAs нуждаются в активации малой GTPase ADP-ribosylation factor (ARF) для ассоциации с мембраной. Соответственно, оба класса адапторов соединяются с ARF также как и с трансмембранным грузом и клатрином (Табл. 1; Рис. 1b). AP-1 γ-субъединица и GGAs несут родственные C-терминальные ‘ear’ домены (γ-ears) которые, подобно α-ears, служат в качестве белок-связывающих платформ (Табл 1). Однако, роль γ-ear связывающих белков в формировании CCV и способ связывания γ-ear неясны. Ведется идентификация новых γ-ear-связывающих ENTH- и ANTH-доменовых белков, участвующих в переносе белков между TGN и эндосомами и характеризуются γ-ear-связывающий мотив и соотв. места связывания на γ-ear доменах.
Mammalian ENTH domain protein at the
TGN/endosomes
Белок млекопитающих KIAA0170 идентифицирован независимо при масс спектрометрии CCVs головного мозга, масс спектрометрии AP-1 γ-ear связывающих партнеров и и в результате поиска в базе данных по ENTH-доменовым белкам. Этот белок был назван enthoprotin, epsinR и Clint; авт. используют обозначение epsinR. Иммунофлюоресцентная локализация и in vitro binding assays показали, что epsinR ассоциирует с элементами TGN и эндосом, которые содержат клатрин, AP-1 и возможно GGA. Неожиданно было обнаружено, что epsinR соединяется с AP-2, а также с AP-1 в экстрактах из головного мозга, хотя распределение epsinR не перекрывается с распределением AP-2. EpsinR строго взаимодействует с γ- и β2-ears, a количественные измерения показалив 40–50-раз более низкое сродство к α-ears. Несмотря на взаимодействие с AP-2 β2-ear in vitro, in vivo оно м. оказаться таковым, что β-ear взаимодействие будет происходить с блихзко родственной AP-1 β1 субъединицей, это позволит epsinR соединяться с обоими AP-1 ear доменами.
EpsinR содержит предположительно ENTH domain signature
мотив и лишен богатого лизином ANTH мотива. Сравнение последовательностей и структурное моделирование epsinR ENTH с epsin1 ENTH обнаруживают различную связывающую специфичность помимо общего сродства к phosphoinositides (Рис. 2c). В самом деле, epsinR ENTH обнаруживает слабое предпочтение к PI(4)P в nitrocellulose
overlay и liposome binding assays. Подобно epsin1 ENTH, epsinR ENTH, по-видимому, играет важную роль в субклеточной локализации epsinR благодаря своей липид-связывающей активности. Трансфекция tagged epsinR ENTH выявляет локализацию, сходную с таковой эндогенного epsinR. Мутации в epsinR ENTH, которые снижают связывание липидов обусловливают изменение распределения трансфицированного полной длины epsinR. Однако, в отличие от сходных epsin1 мутаций, мутантный белок все еще ассоциирован с внутриклеточными мембранами, указывая тем самым на более сложный механизм ассоцииации с мембраной epsinR.
Предсказанная helix 0 у epsinR экспозирует гидрофобную поверхность к epsin1 helix 0 (Рис. 2c). К сожалению, ассоциация epsinR ENTH домена с липосомами не достаточно строга, чтобы м.б. использовать подход liposome tubulation
или monolayer bending. Однако, мутация центрального гидрофобного остатка в helix 0 заставляет трансфицированный белок локализоваться в виде аномально больших точек (puncta), это указывает на то, что гидрофобная природа жтой области вносит вклад в собственно распределение epsinR. Все это говорит о том, что ENTH домен способен направлять psinR в TGN/endosomes и м. облегчать изгиб мембраны во время образования CCV.
EpsinR содержит два clathrin-связывающих мотива (Рис. 1c),
более C-терминальный мотив обеспечивает основную связывающую клатрин активность in vitro. Слабое связывание липидов с помощью epsinR не позволяет использовать подход рекрутирования клатрина в монослой. Тесты на сборку клатриновой решетки с помощью разных фрагментов epsinR дали противоречивые результаты, возможно из-за влияния
N-terminal последовательностей помимо клатрин-связывающих мотивов. Установлено, что epsinR является стабильным компонентом CCVs. В этой связи, epsinR напоминает AP180 больше, чем epsin1, этим открывается возможность, что ассоциация epsinR с клатриновой решеткой создает AP180-подобную структурную поддержку. Функция epsinR в CCV-обеспечиваемом транспорте между TGN и эндосомами тестировалась с помощью избыточной экспрессии и siRNA. Подтверждена роль epsinR в сортировке лизосомных энзимов из TGN, избыточная экспрессия epsinR полной длины обусловлвивает дефектную сортировку cathepsin D в лизосомы и снижение уровня ассоциированных с CCV
mannose 6 phosphate рецепторов, сортинг-рецепторов для cathepsin D. Напротив, siRNA снижение экспрессии epsinR не влияет на сортировку cathepsin D в условиях, при которых siRNA-обусловленное снижение AP-1 вызывает дефекты сортировки. Этот важный результат м.б. обусловлен перекрыванием функции др. белком или участием epsinR в др. пути.
Yeast ENTH/ANTH domain proteins function in
TGN/endosome transport
Специальная статья, посвященная этому вопросу "Yeast epsin-related proteins required for Golgi–endosome traffic define a γ-adaptin ear-binding motif" в виде *.pdf файла находится
Двугибридный скрининг с использованием ear доменов дрожжевых AP-1
γ-субъединицы и Gga2p выявил два белка, Ent3p и Ent5p. Оба белка связывают AP-1 γ и Gga ear домены in vitro и ко-иммунопреципитируются с Gga2p. Ent5p иммунопреципитируется также с AP-1. Ни один не ассоциирует с AP-2. При иммунофлюоресценции Ent3p и Ent5p существенно ко-локализуются с внутренними клатриновыми фокусами, представляющими, как полагают, TGN и эндосомы. Подобно epsinR ENTH, Ent3p N-терминальный домен содержит ENTH signature мотив и м.б. моделирован как amphipathic helix 0 (Рис. 2d). Предсказываемый lipid-связывающий карман Ent3p отличается от такового у epsinR, указывая тем самым, что epsinR и Ent3p м. связывать разные phosphoinositides, хотя оба облегчают изгибаемость мембраны. В отличие от Ent3p, Ent5p несет богатый лизином ANTH-подобный phosphoinositide-связывающий мотив. Т.о., Ent5p вряд ли способствует изгибаемости мембраны и м. функционировать скорее подобно AP180, чем epsin1.
Ent3p и Ent5p, оба необходимы для рекрутирования клатрина in vivo; однако, они отличаются по клатрин-связывающим свойствам. Ent5p имеет два канонических клатрин-связывающих мотива и м.б. ко-иммунопреципитирован с клатрином, тогда как Ent3p лишен клатрин-связываяющих мотивов на своей С-терминальной области и не ко-и ммунопреципитируется с клатрином. Делеция генов, кодирующих и Ent3p и
Ent5p, но не одиночные делеции, обусловливают дефект локализаци клатрина. Неспособность поставлять клатрин на TGN/endosomes сопровождается дефектами клатрин-зависимого переноса между TGN и эндосомами. Это доказывает перекрывание функций ENTH- и ANTH-доменовых белков по сборке клатрина и в CCV-обеспечиваемом транспорте в дрожжевой TGN/endosome системе.
Mapping sites of interaction between γ-ears and
adaptor/accessory proteins
Пролит свет на молекулярную основу распознавания accessory/adaptor
с помощью этих доменов. Первые данные получены из кристаллической структуры AP-1 γ ear, которая представлена 8-нитевым β-sandwich, сходным с одним из двух субдоменов, обнаруженных в большом α-ear. C помощью мутагенеза определили область, связывающую акцессорные факторы, отличную от таковой в α-ear. Остатки в полосе basic остатков и в соседнем неглубоком гидрофобном кармане важны для связывания акцессорного белка, это указывает на то, что соотв. сайты связывания на акцессорных белках м.б. представлены кислыми и гидрофобными остатками. В соответствии с предсказанием EpsinR, Ent3p, Ent5p, γ-synergin и rabaptin-5 содержат законсервированные γ-ear-связывающие мотивы с общими консенсусными последовательностями: [D/E][G/A](0,1)F[G/A][D/E]B
(Рис. 3). Изменчивость последовательностей между мотивами указывает на незначительные отличия в способе связывания. Напр., мотив rabaptin-5 является более гидрофобным, чем таковой в γ-synergin, а мутации в basic полоске у AP-1 γ ear не ингибируют связывание rabaptin-5, но устраняют связывание γ-synergin. Такие отличия м. играть важную роль в иерархии рекрутирования оболочек или в предопределении пространственной ориентации между разными взаимодействующими партнерами в формирующихся CCVs.
Fig. 3. Acidic phenylalanine motifs from g-adaptin and GGA-binding proteins and related unstudied proteins. Underlined sequences were shown to be sufficient for binding to either γ-adaptin or GGA ear domains. In red are residues that, when mutated, affect ear binding. D.m.is Drosophila melanogaster. C34E11.1 and SPC794.11c are uncharacterized Caenorhabditis elegans and Schizosaccharo-myces pombe proteins related to epsinR and Ent3p. At the bottom, a consensus sequence, where B indicates hydrophobic.
Неясно, имеются ли дополнительные взаимодействующие поверхности на γ ears и/или альтернативные мотивы взаимодействия. Eps15, который соединяется с γ-adaptin ear, содержит acidic-phenylalanine-подобную последовательность DGFADF, и сходные последовательности присутствуют в auxilin-2. Напротив, кандидаты в мотивы не столь легко обнаруживаются у двух недавно выявленных γ-ear gfhnythjd, Snx9 и Arf-GAP1.
Concluding remarks
Recent characterizations of epsin- andAP180-like molecules
involved in CCV formation at the TGN/endosomes in
mammalian and yeast cells suggest that these protein
families constitute a fundamental element in nucleation of
clathrin-coated vesicles. However, these new advances in
understanding CCV formation at the TGN and endosomes
raise many important questions. Do epsinR and Ent3p
actually contribute to membrane curvature? Are there
additional ENTH/ANTH domain proteins that cooperate
with epsinR? What are the lipid-bindingspecificities of yeast
Ent3p and Ent5p and how does Ent3p, which
lacks functional clathrin-binding motifs, contribute to
clathrin localization? Are there other g-adaptin and/or
Gga-interacting proteins with functions analogous to
endocytic accessory proteins such as amphiphysin and
synaptojanin and/or are there novel accessory proteins
uniqueto AP-1 and/or GGA-mediated CCVformation? What
are the hierarchies of assembly and the regulatory mech-anisms
that control assembly? With identification of epsinR
and the Ent proteins, and definition of a basis for g-ear
recruitment of accessory factors, the foundations are now in
place to address these issues.
У меня развился микоплазмоз и я немому не то что работать ну и ходить теперь стало тяжело


){kind=link}