Сразу после вступления спермия в ооцит при оплодотворении, его ЯО, не содержащая пор, быстро замещается на оболочку мужского пронуклеуса, которая сходна по составу с таковой женского пронуклеуса. "Изменения оболочек" начинаются с разборки двойной мембраны, так что хроматин спермия входит в соприкосновение с цитоплазмой яйца. Это делает возможным быстрое включение материнских гистонов и других белков хроматина и ведет к экстенсивному разбуханию спермия. Затем срезу же начинается сборка ЯО вокруг хроматина для образования мужского пронуклеуса. Tckb большая часть оболочки пронуклеуса происходит из ER яйца, то специализированные области исходной оболочки спермия сохраняются на его кончике и на ассоциированной с центросомами базе ядра. Эти структуры, как полагают, важны для образования новой ЯО.
У морских ежей и др. видос сборка пронуклеуса происходит в интерфазной цитоплазме, в которой уже присутствует женский пронуклеус и нет необходимости в синтезе новых белков. Annulate lamellae (AL) очевидные кандидаты в качестве источника материала для ЯО в этом случае. AL являются специализированными областями ER, упакованными уже собранными NPCs, они многочисленны в большинстве ооцитов. AL, как полагают, содержат NPCs для быстрого их использования во время быстрых ранних эмбриональных делений. В М фазе AL комплексы пор разбираются, в результате растворенные белки ядреных пор становятся доступными для сборки ЯО в анафазе и телофазе. Напротив, у видов, у которых оплодотворение происходит после мейоза II в интерфазе, AL являются интактными, т.к явялются NPCs женского пронуклеуса. Следовательно, разные механизмы сборки д. функционировать, чтобы сформировать мужской пронуклеус. Простейшей моделью будет, что AL прикрепляется к поверхности хроматина в виде предварительно собранных строительных блоков для ядерной мембраны. Эта модель косенно подтверждается тем фактом, что AL движутся в направлении мужского хроматина вдоль звазд спермия и что nocodazole блокирует образование пронуклеуса в основном препятствуя образованию кластеров AL. В самом деле, после деполимеризации микротрубочек AL остается распределенным по всей цитоплазме, а хроматин спермия окутывается мембранами без пор, которые неспособны сливаться с женским пронуклеусом. Сходным образом в экстрактах яиц Xenopus инсерция NPC в в ЯО, которая собирается вокруг хроматина спермия, м.б. блокирована с помощью nocodazole.
Mutual attraction: pronuclear movement
После образования кластеров AL растущая звездочка спермия захватывает женский пронуклеус и два пронуклеуса быстро сближаются с помощью процесса, обеспечиваемого динеином. Сходный механизм, по-видимому, ответственен за динеином-обеспечиваемое прикрепление микротрубочекк ядрам во время NEBD у фибробластов и при миграции центросом у дрозофилы. Кандидатами на роль обеспечения ЯО/AL взаимодействия являются NPCs, т.к. только они являются компонентами и ЯО и AL. Более того, только эти структуры соединяют наружную ядерную мембрану с ламиной, обеспечивая тем самым достаточную механическую стабильность для движения целого ядра посредством микротубулярных моторов. Напротив, простое прикрепление микротрубочек к наружной ядерной мембране возможно только в результате образования ER трубочек, несмотря на то, что это прикрепление обеспечивается комплексами, занимающими околоядерное пространство.
Pronuclear fusion: how to merge two complete nuclear envelopes
Когда пронуклеусы в яйцах морских ежей приходят в тесное соприкосновение, то их ЯО сливаются. Хотя это слияние не является универсальным для всех ооцитов (напр., пронуклеусы мышей только становятся др. против др. и их хромосомы соединяются с общми митотическим веретеном для первого митотичпеского деления дробления), сходные слияния происходят у большинства видов позвоночных после митозов, во время раннего эмбрионального развития в процессе слияния karyomere. Кариомеры это микроядра, которые формируются покруг каждой хромосомы во время анафазы. Их ЯО содержат поры и поддерживают нуклеоцитоплазматический транспорт и репликацию ДНК. Лишь позднее в репликации эти кариомеры сливаются, образуя одно общее ядро, содержащее все хромосомы. Образование кариомеров, по-видимому, связано с необходимостью быстрого вступления в S фазу быстро делящихся бластомеров.
Топологически слияние двух полных ядер вызывает несколько проблем. Поперечно связанные структуры ЯО с двумя ядерными мембранами, высокая стабильность NPCs и ламин в интерфазе. Следовательно, слияние ядер наиболее вероятно нуждается локальной (или полной) разборке ламины, удалении NPCs с места слияния, в специфическом слиянии наружных мембран и специфическом слиянии внутренних мембран (Рис. 3).
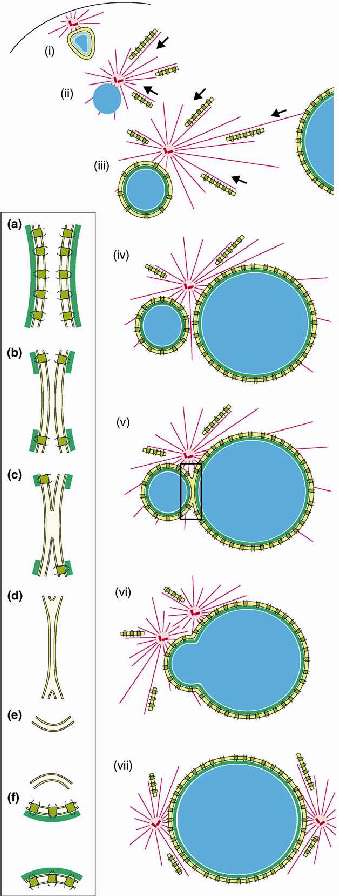 NE dynamics during fertilization in echinoderms. (i) As the sperm enters the oocyte, it is still surrounded by the pore-less sperm NE. (ii) The sperm envelope disassembles and the nucleus swells as a result of chromatin rejrganization. (iii) The pronuclear envelope forms, utilising the NPC reserves stored in the AL, which are moved along the sperm aster microtubules. (iv-vii) Pronuclei then move towards each other and fuse. Insert: A possible model for pronuclear fusion. (A,b) NPCs have to be removed from the fusion site, whoch presumably also requires local disassembly of lamina. (c) Outer membranes then fuse. (D,e) The inner membranes then follow suit. Microtubules and centrosomes, red; chromatin, blue; lamina, green; ТУб yellow; NPCs, dark yellow.
NE dynamics during fertilization in echinoderms. (i) As the sperm enters the oocyte, it is still surrounded by the pore-less sperm NE. (ii) The sperm envelope disassembles and the nucleus swells as a result of chromatin rejrganization. (iii) The pronuclear envelope forms, utilising the NPC reserves stored in the AL, which are moved along the sperm aster microtubules. (iv-vii) Pronuclei then move towards each other and fuse. Insert: A possible model for pronuclear fusion. (A,b) NPCs have to be removed from the fusion site, whoch presumably also requires local disassembly of lamina. (c) Outer membranes then fuse. (D,e) The inner membranes then follow suit. Microtubules and centrosomes, red; chromatin, blue; lamina, green; ТУб yellow; NPCs, dark yellow.
Рассасывание ламины скорее всего является первой ступенью, т.к. оно делает возможным диффундироваль латерально закрепленным NPCs. Как только NPCs уходят из места слияния, то наружные мембраны м. сливаться, образуя промежуточное состояние с двумя внутренними мембранами, лежащими в тесной близи. Сходные промежуточные состояния видны на ЭМ, укахывая тем самым, что таки структуры существуют в течение продолжительного периода времени. Затем внутренние мембраны также сливаются, содержимое двух ядер смешивается. Первоначально только узкий канал соединяет два прнуклеуса, который медленно увеличивается в размерах. Очевидно, что слияние пронуклеусов нуждается в хорошо действующей machinery. Оно м. иметь общие свойства с ремоделированием интерфазных ядер во время репликации и вирусной инфекции.

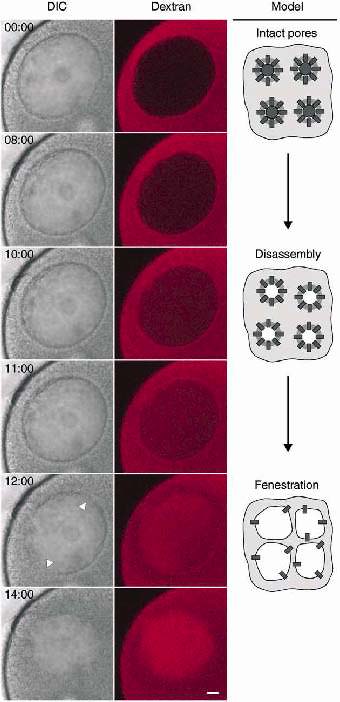 Dextran entry into the nucleus in maturing starfish oocytes: the two phases of NEBD. Tetramethyl-rhodamine-labeled 70 kDa dextran was injected into the cytoplasm of the oocyte/ Before any change could be seen on the differential interference contrast (DIC) image, the dextran slowly starts to enter the nucleus (frames 8:00-11:00), reflecting the beginning of the disassembly of the pore complex. The slow entray is then followed by a rapid wave of dextran entray (frame 12:00), coinciding with the disappearance of the sharp nucleoplasmic boundary on the tramsmitted light image (arrowheads). Time is given as minutes:seconds. Bar =10 µm. The scheme illustrates the 'top view' of the NE. Model is adapted from [19].
Dextran entry into the nucleus in maturing starfish oocytes: the two phases of NEBD. Tetramethyl-rhodamine-labeled 70 kDa dextran was injected into the cytoplasm of the oocyte/ Before any change could be seen on the differential interference contrast (DIC) image, the dextran slowly starts to enter the nucleus (frames 8:00-11:00), reflecting the beginning of the disassembly of the pore complex. The slow entray is then followed by a rapid wave of dextran entray (frame 12:00), coinciding with the disappearance of the sharp nucleoplasmic boundary on the tramsmitted light image (arrowheads). Time is given as minutes:seconds. Bar =10 µm. The scheme illustrates the 'top view' of the NE. Model is adapted from [19].