Zebrafish foxi one modulates cellular responses to Fgf signaling required for the integrity of ear and jaw patterning Development 130, 2543-2554, 2003 | |
|
Идентифицировано 4 инсерционных аллеля foxi one (foo), эмбриональной летальной мутации у рыбок данио, которые обнаруживают дефекты как отической плакоды, так и челюсти. У эмбрионов foo/foo отическая плакода расщепляется на две маленькие плакоды и мутантны обнаруживают дефект dorsoventral (DV) хряща, проявляющиеся как редуцированная hyomandibular и редуцированные третья и четвертая бранхиальные дуги. Идентифицированый foxi one (foo), это ортолог у рыбок данио Foxi1 (FREAC6, FKHL10, HFH-3, Fkh10) и член семейства forkhead доменовых регуляторов транскрипции, т.к. этот ген мутантен у эмбрионов foo/foo. Ген foo экспрессируется в клетках предшественника, a foo/foo эмбрионы лишены экспрессии pax8 в плакоде и имеют дизорганизованную отическую экспрессию pax2.1 и dlx3.
Третий поток миграции клеток нейрального гребня, выявляемый с помощью экспрессии dlx2 и krox20, является аберрантным, т.к. он проникает на территорию отической плакоды. foo экспрессируется в энтодерме глоточного кармана и необходим для экспрессии в кармане pax8 и собственно для формирования паттерна др. маркеров в кармане, таких как nkx2.3. У эмбрионов foo/foo не поддерживается экспрессия fgf3 в карманах, это сопровождается апоптозом клеток нейрального гребня в соседней дуге. Предполагается, что экспрессия foo обязательна для экспрессии pax8 и возможно стоит ниже Fgf signaling в законсервированном пути совместно необходимому для интеграции формирования паттерна отической плакоды и глоточного кармана. Предполагается, что правильное помещение survival/proliferation сигналов существенно для формообразования фарингеальных хрящей и что эволюционная связь между формированием челюстей и уха м. б. прослежена с помощью Fgf-Foxi1-Pax8 пути.
| |
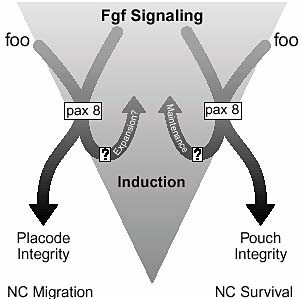 Fig. 10. A model that places foo in pathways for ear and jaw
development. foo expression is independent of Fgf signaling, but it
allows cells to respond to Fgf signaling with a specific genetic
program, which includes the induction of pax8. Feedback
mechanisms for the maintenance of Fgf expression are inferred from
the data. Phenotypic effects observed in the embryos when foo
function is eliminated are listed below.
Fig. 10. A model that places foo in pathways for ear and jaw
development. foo expression is independent of Fgf signaling, but it
allows cells to respond to Fgf signaling with a specific genetic
program, which includes the induction of pax8. Feedback
mechanisms for the maintenance of Fgf expression are inferred from
the data. Phenotypic effects observed in the embryos when foo
function is eliminated are listed below.