Self-organization and forces in the microtubule cytoskeleton Current Opinion in Cell Biol. V. 15. P. 118-124 | |
|
|
В биологиии термин "само-организация" имеет разные смыслы. Чаще всего имеется в виду процесс организации, в котором множественные агенты следуют правилам поведения, предписанными локальной информацией. Самоорганизация нуждается в отсутствии заранее составленного знания о конечной организации - плана, которому будут следовать агенты. Термин "самоорганизация" используется вместо синонима out-of-equilibrium организация. В этом более точном смысле самоорганизовнные системы постоянно consumes и dissipates энергию для поддержания самих себя. Это д.б. противопоставлено "само-собирающимся" системам, которые вместо высвобождения свободной энергии во время своей организации, ведут к статичным структурам, в которых нет тока энергии. Само-сборка и само-организация, т. о., покрывают две основные возможности в отношении потребления энергии. Обычно феномен организации включает в себя и само-сборку и само-организацию.
The cytoskeleton as a self-organising system Цитоскелет основа внутренней архртектуры клетки и его организация лучше всего объясняется самоорганизацией. Это касается ключевых компонентов цитосклета: полярных филамент, генерируемых с помощью нековалентной сборки тубулиновых или актиновых субъединиц. Эта сборка связана с ГТФ и АТФ гидролизом, для микротрубочек и актина, соотв. Динамические филаменты, обнаруживаемые в клетках, являются само-организовывающимися структурами, т.к. они постоянно потребляют энергию. Микротрубочки или актиновые филаменты м.б. также обозначены и как само-собирающиеся структуры, т.к. он м. формироваться даже при ингибировании гидролиза ГТФ - это происходит без существенного потребления энергии. Следовательно, и само-сборка и само-организация м. давать микротрубочки; однако, два типа микротрубочек имеют разную природу. Их тубулиновые решетки хотя и сходны, выглядят по-разному на электронных микрофотограциях. Эти различия будут еще отчетливей, если выявить силы и натяжение внутри структуры. В то время как статические микротрубочки, растущие в присутствии неспособного к гидролизации ГТФ аналога, являются трубочками, которые обнаруживают малое натяжение, то динамические микротрубочки, растущие в присутствии ГТФ, являются трубочками легко разрушаемыми. Во время роста энергия, поставляемая за счет гидролиза ГТФ, хранится в решетке в виде механических натяжений (strain). Эти нятяжения создают силы быстрого укорочения разбирающихся микротрубочек. Силы нелегко видеть и лучшим способом выявления само-организации является выявление динамических свойств системы. Динамическая нестабильность и однообразная механическая работа (treadmilling) являются феноменом, который нуждается в рассеивании энергии и который не встеречается в процессах чистой само-сборки.
Т.о., тубулиновые или актиновые мономеры само-организуются в динамические филаменты. Следущий уровень сложности касается саморганизации этих филамент в различные трехмерные паттерны. Для этого необходимы др. компоненты, такие как молекулярные моторы. В проивоположность химическим системам, таким как реакции Белоусова-Жаботинского, механические силы являются центраьымими в самоорганизации цитосклета, просто из-за размеров этих филаментов. В самом деле, в то время как малые молекулы м. диффундировать и быстро реагировать, чтобы огранизовать самих себя в пространстве, диффузия цитоскелетных филаментов слишком мала, чтобы играть важную роль в их организации на временной шкале жизни клетки. Различные механизмы используются, чтобы достичь этой медленной диффузии. Одним из таких механизмов является направленный транспорт филамент с использованием, напр.. механических сил, генерируемых молекулярными моторами, которые потребляют энергию в форме гидролиза АТФ. Кроме того, ограничение, граыитация, градиенты концентраций мономеров и стерические взаимодействия меэжу филаментами вносят вклад в самоорганизацию филамент. Др. механизмом является возникновение (nucleation) филамент непосредственно в соответствующем месте. Др. стратегии используют динамические свойства концов филамент. Пространственая регуляция динамической нестабильности микротрубочек также м. б. результатом разборки филамент с нежалательными характеристиками. Необходимо определить относительный вклад нуклеации, транспорта и динамической стабилизации в генерацию наблюдаемых паттернов.
Pathways of aster formation В клетках животных микротрубочки часто организовываются в форме звезд, которые своими плюс концами расходятся в направлении периферии плетки, а их минус-концы сфокусированы вблизи ядра. Одним из механизов, ведущих к такой ораганизации, является локализованная нуклеация (зарожедение): тубулин не м. собираться спонтанно в цитоплазме, поэтому центросома предопределяет источник роста микротрубочек (Рис. 1а).
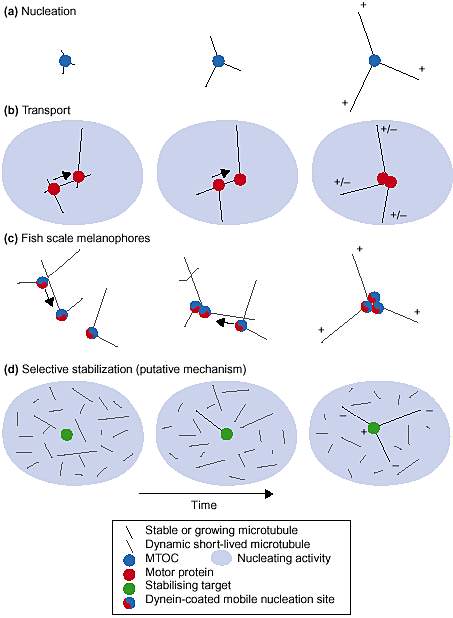 Polar radial arrays of microtubules - asters - can be formed in different ways. (a) Nucleation by a microtubule-organising centre, followed by growth at the plus end. (b) Nucleation throughout the sample, followed by motor-dependet reorganization. Be moving on two mivrotubule simultaneously oligomeric motors organise the microtubules in space. Homo-complexes made of minus-end-directed motors produce asters with normal polarity, while complexes of plus-end-directed motors produce asters of opposite polaruty, with the minus ends out. (c) Nucleation by pigment granules, wich are transported along mivrotubules by the motor dynein. Granules aggregate at microtubule minus ends, while dynamic microtubules disassemble in regions free of granules. (d) Apart frpm these processes that have been observed in living matter, either under natural or forced conditions, one can imagine other pathways leading to asters. For exemple, pure selective stabilization on a spot would lead to the generation of inverted radial arrays with plus ends in the crntre. Indeed, let's assume that we create a cytoplasm in wich short-lived microtubules are nucleated evwrywhere and that there are pointed spots of stabilization distributed in the cytoplasm. All the plus ends that do not hit a stabilising spot will eventually disappear, resulting in the formation of 'inverted asters'.
Polar radial arrays of microtubules - asters - can be formed in different ways. (a) Nucleation by a microtubule-organising centre, followed by growth at the plus end. (b) Nucleation throughout the sample, followed by motor-dependet reorganization. Be moving on two mivrotubule simultaneously oligomeric motors organise the microtubules in space. Homo-complexes made of minus-end-directed motors produce asters with normal polarity, while complexes of plus-end-directed motors produce asters of opposite polaruty, with the minus ends out. (c) Nucleation by pigment granules, wich are transported along mivrotubules by the motor dynein. Granules aggregate at microtubule minus ends, while dynamic microtubules disassemble in regions free of granules. (d) Apart frpm these processes that have been observed in living matter, either under natural or forced conditions, one can imagine other pathways leading to asters. For exemple, pure selective stabilization on a spot would lead to the generation of inverted radial arrays with plus ends in the crntre. Indeed, let's assume that we create a cytoplasm in wich short-lived microtubules are nucleated evwrywhere and that there are pointed spots of stabilization distributed in the cytoplasm. All the plus ends that do not hit a stabilising spot will eventually disappear, resulting in the formation of 'inverted asters'. Это ествественно ведет к генерации радиального паттерна с униформной полярностью. Фактически, по крайней мере, существуют еще два др. пути (Рис. 1). Один из этих путей обнаружен при изучении образования звездочек в экстрактах митотических яиц Xenopus, подвергнутых действию стабилизирующего микротрубочки taxol. В этих экстрактах таксол индуцирует случаную сборку микротрубочек по всей цитоплазме, которые реорганизуются в звеждочки под действием мотора динеина (Рис. 1b). Третьий путь выявлен во фрагментах меланофоров из чешуек рыб (Рис. 1с). Хотя первоначально считалось, что эти звездочки также образуются под действием моторов, связывающих некоторые филаменты, однако то, что микротрубочки были неподвижны, привело авт. к новой гипотезе, бозирующейся на транспорте локальных нуклеаторов (nucleators). В этих клетоных фрагментах пигментные гранулы, по-видиому, нуклеируют микротрубочки. Т.к. гранулы покрыты динеином, то они транспортируются к минус концам микротрубочек. Это создает позитивную петлю обратной связи для локальной агрегации гранул и конкурентной нуклеации микротрубочек, что и ведет к образовнию звездочек. В клеточных фрагментах конкуренция за гранулы между множественными звездочками в конечном итоге ведет к объединению всех гранул вместе. Микротрубочки вскоре деполимеризуются в областях, лишенных гранул, это ведет к образованию одной большой звезды. Образование звезд из центросомы является следствием точного протстранвенного контроля нуклеации. Образование звездочек с помощью олигомерных моторов указывает на направленный транспорт филамент с помощью моторов в некотролируемых условиях нуклеации. Образование звезд в меланофорах рыб представляет собой интересную смесь странспорта и нуклеации. В то время как образование звезды из центросомы происходит за счет чистой само-сборки (с нединамическими филаментами), два др. примера образования звездочек являются настоящими феноменами само-организации. Имеется, пока еще не совсем удовлетворительная аналитическая теория, которая м. предсказывать правильные исходы моторами-обусловленной само-организации исходя из множества кинетических скоростей, описывающих взаимодействия компонентов. Более легкие количественные подходы используются для верификации интуитивных моделей.
Characteristic features of the spindle Как же происходит процесс самоорганизации в митотическом веретене. Во время деления клетки все цитоплазматические микротрубочки обычно исчезают, замещаются митотическим веретеном, которое собирается для физического разделения хромосом. Биполярность необходима не как средство фокусировки двух полюсов, а как средство организации микротрубочек в веретено с четкой ориентацией, вдоль которой и будет происхдить расхождение хромосом. Важным свойством веретена является amphitelic прикрепление кинетохор, которое необходимо для растаскивания сестринских хроматид в противоположных направлениях. Более того, силы, прикладываемые к кинетохорам, д. иметь тенденцию стянуть два полюса вместе, если бы не коменсировались действием противоположных сил, возникающих или в результате физического соединения полюсов - спсобного противостоять компрессии - или в результате соединения полюсов с ригидным клеточным кортексом посредством астральных микротрубочек. Соединение полюсов обычно реплизуется с помощью центрального антипараллельного перекрывания микрорубочек, возникающих на каждом из полюсов, которые м. увидеть непосредственно после начала анафазы. Второй признак, который м.б. не существенным для всех клеток, связан с наличием хорошо сфокусированых полюсов, соотв. расположенных хроматид или кинетохоров, дающих узкую метафазную пластинку. Напр., веретена в царстве животных имеют свои полюса хорошо сфокусированные с помощью центросом, тогда как в царстве растений имеются широкие полюса.
Веретено м.б. , следовательно,описано как комбинация простых геометрических характеристик. Большинство из этих призанков является до некоторой степени избычтоным и перекрывающимся. М. ожидать, напр., что фокусировка полюсов с помощью двух центросом распространяет биполярность на структуру. Сходным образом, антипараллельное перекрывание микротрубочек в срединной зоне веретена также создает биполяризацию структуры. Однако подсистемы веретена м.б. подразделены. Используя систему покрытых хроматином кусочков, удалось показать, что формирование полюсов в отсутствие центросом происходит в процессе, очень сходном с мотор-зависимым образованием звездочек. На живых клетках позвоночных подтверждено, что биполярное веретно м. формироваться в отсутствие центросом по способу, напоминающему таковой в растительных клетках. Напротив, центросомами нуклеированные звездочки м. взаимодействовать в отсутствие хроматина: напр., у морского ежа, в лизатах Xenopus, у гомокарионов, генерируемых при слиянии PrK (почек кенгуру) клеток, или в энуклеированных сперматоцитах. В бесхромосомных ооцитах мышей илия экстрактах яиц Xenopus при добавлении постоянно активной мутантной формы регуляторного белка Ran, формируются структуры микротрубочек. Хотя они лишены и хромосом и ценромер, эти структуры микротрубочек имеют биполярную организацию и возможно также антипараллельные взаимодействия - два признака, характерные для веретена. Даже смеси очищенных моторов и микротрубочек м. продуцировать само-органиозванные структуры, с некоторыми характеристиками веретена, напр., звездочки или разделение моторов, направленных на плюс- и минус-концы.
Pathways of spindle assembly Предложено множество гипотез для объяснения морфологии веретена. Напр., захват и селективная стабилизация центросомами-нуклеированных микротрубочек с помощью кинетохор считается общим механизмом. Эта "поиск-и-захват" схема и в самом деле м. объяснить, как кинетохоры прикрепляются к микротрубочкам веретена, но не м. объяснить биполярность в экстрактах Xenopus вокруг покрытых хроматином кусочков в отсутствие центросом и кинетохор. Чтобы объяснить эти фигуры было предположено, что нуклеация локальных микротрубочек и стабилизация вокруг хромосом связаны с их организацией в биполярное расположение с помощью сложной смеси моторов, направленных к плюс- и минус-концам. Эта модель объясняет биполярность, но не м. объяснить образование прикрепления специфических микротрубочек к кинетохорам, процесс важный для расхождения хромосом.
Эти две идеи - также как и экспериментальные системы или спровоцированные - действительно адрессованы разным геометричесикм свойствам веретена: поиск-и-захват объяснеет прикрепление кинетохор; тогда как мотором-обусловленная самоорганизация является главным объяснением биполярности. Необходимо комбинировать эти идеи ( и др.) в полной модели на принцыпах организации филамент (Рис. 1). Модель сборки веретена вокруг хроматиновых кусочков является комбинацией нуклеации и моторами-обеспечиваемого транспорта. Нуклеация не ограничена небольшой областью хромосом, а образует галдо вокруг хроматина, создаваемое за счет диффузнного градиента еще неизвестных эффектов. Известно, что градиент существует и локально активируется с помощью Ran GTP. Фактически этот градиент м. влиять как локальную нуклеацию микротрубочек, динамику микротрубочек и их плюс-концы, а также активность моторов. Во всяком солучае модель строится на тех же трех принципах, но с др. пространственными регуляциями, точками или градиентами (Рис. 2).
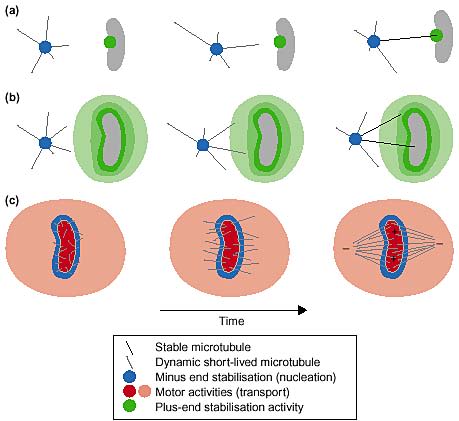 Proposed models of spindle morphogenesis are combinations of nucleation, motor activity and microtubule stabilisation. (a) Search-and-capture combines localised nucleation with localised stabilization. (b) Chromosomes could stabilise microtubules a-distance, through an as yet unidentified diffusible factor. (c) Chromosomes can also induce the nucleation of microtubules in their vicinity. Together with molecular motors, they would organise into a focused bipolar array.
Proposed models of spindle morphogenesis are combinations of nucleation, motor activity and microtubule stabilisation. (a) Search-and-capture combines localised nucleation with localised stabilization. (b) Chromosomes could stabilise microtubules a-distance, through an as yet unidentified diffusible factor. (c) Chromosomes can also induce the nucleation of microtubules in their vicinity. Together with molecular motors, they would organise into a focused bipolar array.
Каждая геометрическая характеристика веретена (биполярность, сфокусированные полюса, антипараллельное перекрывание и т.д.) м.б. сложной проблемой. Данные по образованию звезд указывают на то, что данный паттерн м. б. получен разными путями самоорганизации. Комбинирование таких путей м. позволить большое количество формальных возможностей образования веретено-образных форм. Многие из этих комбинаций будет приводить к структурам, напоминающим веретено, но неспособными обеспечивать сегрегацию хромосом. Их структура м.б. несовместима с анафазой, напр., они м.б. неспособными продуцировать достаточное количество тянущих сил на кинетохоры, чтобы их разделить. Фактически, генерация этих сил необходима для веретена, чтобы стать растяжимой (tensile) структурой, когда оно достигает стабильного состояния в метафазе. Существование таких натяжений продемонстрировано с помощью лазерной микрохирургии на эмбрионоах Caenorhabditis elegans. Повреждения в середине веретена приводили к разделению двух половин, которое происходило значительно быстрее, чем разделение хромосом в анафазе. Хотя техника не позволяет измерить количественно эти силы, но результаты указывают на то, что веретено находится под значительно большим натяжением, чем это необходимо для растаскивания хромосом. Эти натяжения, как полагают, возникают в результате антогонистического действия большого числа моторных белков, чей химический цикл продуцирует силы и которые также чувствительны к силам в структуре, которые м. замедлять их угасание. Рост микротрубочек м. б. также чувствителен к противостоящим силам. Неожиданно, натяжения на кинетохоры используются также для передачи сигналов - они являются источником каскада регуляционных реакций, запускающих в результате митотические checkpoint - позволяя митозу перейти в анафазу. Следовательно, морфогенез митотического веретена является процесом самоорганизации, который создает очень точную форму и свои собственные внутренние напряжения, чтобы тянуть кинетохоры.
|