В конечностях, как и в сомитах, онтогенетические программы мышц и сухожилий тесно связаны, но имеется важное отличие вих обоюдной зависимости. Т.к. мышцы и сухожилия в проксимальных частях конечностей формируются в тесном контакте др. с др., а популяции их предшественков м. первоначально развиваться автономно, даже проходя поздние морфогенетические события, такие как подразделение мышечных масс и индивидуализация зачатков сухожилий, необходимо, чтобы они взаимодействовали. Т.о., в отличие от сомитов, где по-видимому, действительно индукция и дифференцировка сомитных предшественников сухожилий зависит от от присутствия дифференцированного миотома, формирование происходящих из латеральной пластинки предшественников сухожилий прооисходит независимо от мышц - даже в ходе развития зрелых, функциональных сухожилий необходима реципрокность между ними.
Что общего между двумя популяциями предшественников сухожилий, так это их взаимоотношения с предшественниками скелета. Наши находки, что сомитные предшественники сухожилий происходят из склеротома, указываею на то, что сухожилия и хращи имеют общего предшественника. Особый интерес вызывает вопрос, когда во время развития сомитов эти два клона дивергируют.Мы наблюдали. что на стадии XVI сомитов у 3.5 дн. эмбрионов экспрессия Scx активируется в субпопуляции склеротома. Интересны исследования, при которых участки склеротома пересаживали в дорсальные сигнальыне условия, показавшие, что склеротом не детерминирован к судьбе хряща вплоть до ст. XII у 3.5 дн. эмбрионов, это указывает на то, что возникновение клонов сомитного храща и сухожилия происходит примерно в то же самое время. Напротив, миотом детерминируется значительно раньше. Имеются сходные общие предшественники для клонов хрящей и сухожилий для конечностей. Так, мышцы конечностей, пекторальные и абдоминальные имеют сомитное происхождение, а ассоциированные сухожильные и хрящевые элементы, которые возникают их мезодермы латеральных пластинок, по-видимому, подобны их сомитным аналогам и связаны довлольно тесно. Хотя неясно, когда эти клоны латеральной пластинки дивергируют, имеются некоторые указания на то, что в конечностях судьба хряща детерминаируется меэжду стадиями 21-23, это примерно соответствует тому времени, как и в сомитах, когда в конечностях инициируется экспрессия Scx.
После момента появления общих предшественников для сухожилий и хрящей в сомитах и латеральной пластинке, д. ли в них появиться сходные сигналы для индукции судьбы сухожильных предшетвенников? Анализ развития сухожилий конечностей покзал, что индукция
Scx зависит от сигналов из зачатка эктодермы конечности, тогда как ограничение Scx-экспресирующих предшественников сухожилий внутри мезенхимы зависит от активности ВМР, которая м. продиводействовать Noggin. Но ни эктодермальная, ни BMP/Nogin передача сигналов не влияет на индукцию сомитного Scx, однако как и в сомитах наблюдается то, что избыточная экспрессия FGFs в мезенхиме конечностей м. запускать в ней эктопическую экспрессию Scx и Tenascin. FGF4, экспрессирующийся в мышцах, соседствующих с местами будущих мышечно-сухожильных соединений, м.б. ответственен за позитивную регуляцию Scx в конечностях во время последующих мышце-сухожильных взаимодействий. Передача сигналов FGF м. однако играть также роль в действительной индукции предшественников сухожилий конечностей, на это указывают наблюдения, что имплантация смоченных FGF4 кусочков в мезенхиму конечностей до экспрессии Scx вызывают эктопическую экспресиию Scx спустя 4 ч; т.о., ещё не идентифицировнный FGF м. участвовать в индукции Scx. В любом случае, выявляется законсервированная роль FGFs во время развития сухожилий, происходящих и из сомитов и латеральной пластинки. Это указывает на то, что FGFs м. индуцировать экспрессиию Scx во всей мезенхиме, компетентной к восприятию судьбы сухожилий.
Scx, thr Syndetome, and Tendon Development
Итак прослежено появление нового сомитного компартмента синдетома внутри склеротома в ответ на сигналы от миотома. Показана постоянная экспрессия Scx во время развития аксиальных сухожилий из популяции предшественников. Идентифицирован путь передачи сигналов миотомного FGF, который действует и индуцируя экспрессию Scx в синдетоме и подразделяя склеротом на Рах1-экспрессирующие предшественники хряща и на Scx-экспресирующие предшественники сухожилий. Синдетом м.б. фактически одним из серии сомитных субкомпартментов, предназначенным для продукции пула предшественников для специфического типа клеток. Синдетом возникает в результате взаимодействия между предшественниками мышц (миотом) и хрящей (склеротом), помещая при этом зачаток сухожилий не только в месте их взаимодействия, но и в том расположении, которое в последующем будет необходимо для выполнения роли связующего звена мышц с хрящами.
Сайт создан в системе
uCoz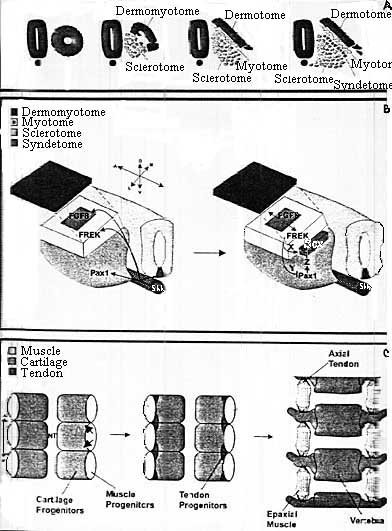 Model for Syndetome Formation
Model for Syndetome Formation