Мы установили, что отклонения в передаче сигналов Notch в параксиальной мезодерме эмбрионов мышей меняют качественные особенности позвонков. Полученные результаты предоставляют прямые доказательства того, что координация сегментации и позиционная спецификация тканей, производных мезодермы вдоль передне-задней оси тела, нуждаются как в достаточных уровнях сигналов Notch, так и циклической активности Lfng.
Позвоночный столб msd::Dll1
dn трансгенных мышей обнаруживает
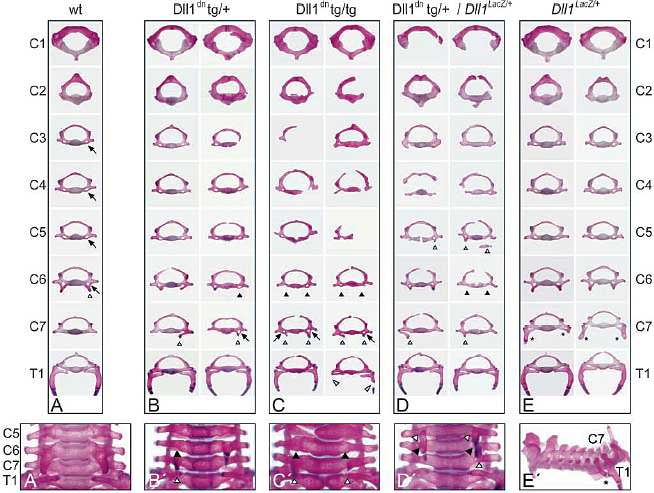
незначительные дефекты, которые, по-видимому, отражают последствия нарушений полярности сомитов. В основном нормальный позвоночный столб, позволяет однозначно идентифицировать отсутствие или эктопическое присутствие характерных признаков (напр., anterior tuberculi и transverse foramina), характерных для определенных позвонков. Потеря таких признаков из некоторых позвонков и их эктопическое появление на др. указывает на изменения качественных особенностей позвонков и указывает на гомеозисные трансформации. Изменения качественных особенностей позвонков обнаружены также у мышей, экспрессирующих Dll1
dn под контролем промотора Mesp2. У таких мышей не выявлено дополнительных уродств позвонков, это подтверждает, что изменения качественных особенностей позвонков у msd::Dll1
dn мышей возникают независимо от дефектов, связанных с сегментной полярностью. Отсутствие дефектов позвонков показательно для нарушенного передне-заднего паттерна у Mesp2::Dll1
dn мышей, это удивительно, т.к. активность Notch в передней части psm является критической при компартментализации сомитов i (Takahashi et al., 2003; Takahashi et al., 2000). Разумным объяснением м.б. то, что активность Notch у Mesp2::Dll1
dn трансгенных эмбрионов выше, чем у msd::Dll1
dn эмбрионов, это подтверждается остаточной экспрессией Hes5 (Fig. 5D), и этого достаточно для установления сегментной полярности. Элемент msd управляет экспрессией мРНК в задней части psm и во вновь образующихся сомитах, но его экспрессия слабая или отсутствует в передней области psm, соотвествующей S-I/S0 (Beckers et al., 2000), тогда как промотор Mesp2 управляет экспрессией специфически в этой области (Fig. 5B). Т.о., в дополнение к кажущейся строгой редукции активности Notch клетки параксиальной мезодермы эмбрионов msd::Dll1
dn подвергаются воздействию редуцированной активности Notch во время большей части её прогрессии через psm, тогда как у Mesp2::Dll1
dn эмбрионов активность Notch редуцирована в клетках только незадолго перед образованием сомита, это и м. вносить вклад в фенотипические различия. Формально нельзя исключить того, что экспрессия Dll1
dn во вновь формируемых сомитах вносит вклад в изменения качественных особенностей позвонков. Однако, это кажется мало вероятным, т.к. экспрессия в передней части psm у Mesp2::Dll1
dn эмбрионов достаточна для воздействия на качественные особенности позвонков. Кажущееся ограничение изменений качественных особенностей позвонков шейной и верхней торакальной областью у msd::Dll1
dn и гетерозиготных Dll1
lacZ мышей м. отражать более высокую чувствительность передних Hox генов к снижению активности Notch.
Шейная область трансгенных мышей, экспрессирующих Dll1
dn, обнаруживает передние трансформации, тогда как гетерозиготные Dll1
lacZ мыши, которые преимущественно имеют редуцированные Dll1-обусловленные сигналы, обнаруживают задние или двунаправленные трансформации, а Dll1
dn мыши, лишенные только одной копии Dll1 имеют смешанный фенотип. Dl1
dn представленный клетками сетчатки цыплят глух к восприятию сигналов Notch (Henrique et al., 1997), блокирование активности Notch осуществляется клеточно автономно. Т.о, Dll1
dn, по-видимому, не только редуцирует Dll1-обусловленные Notch сигналы, но и м. также влиять на сигналы, обеспечиваемые др. лигандами и разными рецепторами, это м. приводить к разным исходам передачи сигналов Notch в psm по сравнению с редукцией только Dll1. Это м.б. не только количественным эффектом, т.к. и у Dll1
dn и у Dll1
dn/Dll1
lacZ/+ эмбрионов экспрессия Hes5 тяжело подавляется и перестаёт обнаруживаться (Fig. 3 and data not shown). Такой способ действия доминантно-негативного Dll1 м. объяснить разные фенотипы у Dll1
dn и Dll1
dn/Dll1
lacZ/+ мышей и м.б. подразумевать, что сигналы, обусловленные разными лигандами или рецепторами вносят вклад в позиционную спецификацию и м. действовать потенциально противоположными путями, сходными с не перекрывающимися или даже противодействующими функциями во время компартментализации сомитов (Takahashi et al., 2003).
Исходя из тяжелой редукции экспрессии Hox генов на 8.5 RBPj k мутантных эмбрионов (Zakany et al., 2001), которые лишены активности Notch, м. ожидать, что ослабленная передача сигналов Notch ведет к редукции экспрессии Hox. Результаты анализа экспрессии 15 Hox генов с помощью in situ гибридизации у
Dll1
dn эмбрионов между 8.5 и 10.5 днём не подтвердили эту идею, хотя нельзя исключить слабый уровень различий, который не выявляется при анализе. Однако границы передней экспрессии Hoxb6 и Hoxc5 были сдвинуты, это указывает на то, что точное позиционирование ростральной границы экспрессии Hox нуждается в точно регулируемой активности Notch в psm. В целом активация Hox в параксиальной мезодерме, по-видимому, серьёзно затрагивается только если передача сигналов Notch сильно снижена или полностью блокирована потенциально уже в предшественниках параксиальной мезодермы. Как активность Notch и регуляция транскрипции Hox генов купированы молекулярно, неясно. Трансформации качественных особенностей позвонков, передний сдвиг экспрессии Hoxb6 и позиции зачатков передних и задних конечностей, обнаруживаются у мышей с отсутствием функции Lfng или при конституитивной экспрессии Lfng. Видимый передний сдвиг экспрессии Hoxb6 у Lfng мутантных эмбрионов м.б. также ожидаем, если меньше сегментов генерируется в проспективной шейной области, тогда как абсолютная позиция границы передней экспрессии Hoxb6 вдоль передне-задней оси тела сохраняется. Недавние модели сегментации сомитов указывают на то, что взаимодействие градированного распределения FGF (Dubrulle et al., 2001) или WNT (Aulehla et al., 2003) сигналов с сегментационными часами генерирует периодический паттерн сомитов. Концептуально, увеличение крутизны градиента или снижение периодичности часов д. приводить к образованию меньшего числа сегментов, которые, во всяком случае, д. становиться больше. Т.о., если потеря Lfng будет затрагивать часы (output) и будет формироваться меньше сегментов в проспективной шейной области, то они д.б. крупнее нормальных. Однако 5 шейных сегментов у Lfng мутантных эмбрионов занимают по существу то же самое пространство, как и передние 5 сегментов WT эмбрионов (Fig. 8), это строго подтверждает, что граница ростральной экспрессии Hoxb6 в самом деле сдвинута кпереди. Положение зачатков передних и задних конечностей инвариантны у эмбрионов дикого типа и соответствуют переходам между шейной и торакальной и поясничной и крестцовой областями соотв. (Burke, 2000). Их передний сдвиг подтверждает гомеозисные трансформации во всём туловище вдоль передне-задней оси тела, это ведет к общей редукции количества сегментов в туловище.
Эксперименты на эмбрионах кур показали, что клетки psm становятся детерминированными в отношении программы сегментации и экспрессии Hox генов в передней трети psm на уровне, обозначаемом как 'determination front', который, по-видимому, предоставляет пороговый уровень FGF8 (Dubrulle et al., 2001).
Расширенное воздействие на клетки FGF8 в передней части psm эмбрионов кур нарушало положение сомитных границ и сдвигало границы экспрессии Нох генов кпереди (Dubrulle et al., 2001), a hypo- и hypermorphic мутации в FGFR1 вызывали гомеозисные трансформации и лёгкий сдвиг грниц экспрессии Hox генов у эмбрионов мыши (Partanen et al., 1998), указывая тем самым, что передача сигналов FGF в передней части psm играет критическую роль в позиционировании границ экспрессии Hox. У эмбрионов мышей транскрипционный взрыв некоторых Hox генов в передней части psm коррелирует с циклической экспрессией Lfng, это ведет к идее, что транскрипционная регуляция Hox генов непосредственно перед образованием сомита происходит в ответ на циклический исход активности Notch, это м. купировать сегментацию с приобретением аксиальных качественных особенностей (Zakany et al., 2001). Трансформации качественных особенностей позвонков вдоль передне-задней оси тела у мышей без функции Lfng также как и при нециклической экспрессии Lfng в psm предоставляет прямые экспериментальные доказательства того, что циклическая активность Lfng существенна для скоординированной генерации сегментов с их позиционной спецификацией. Т.к. в целом спецификация разных анатомических регионов сохраняется, то колинеарность экспрессии Hox генов наиболее вероятно не затронута. Было предположено (Dubrulle et al., 2001; Zakany et al., 2001), что достижение точной комбинаторной экспрессии генов Hox в сомитах происходит в две ступени: во-первых, наиболее вероятно в предшественниках параксиальной мезодермы перед их вступлением в psm кластеры Hox д. прогрессивно открываться и становиться psm, и дефинитивная экспрессия генов Hox д.б. распределена по сегментным единицам вкупе с сегментационными часами. Наши результаты согласуются с критической ролью передачи сигналов Notch и циклической активностью Lfng на второй ступени, потенциально после того, как клетки пройдут фронт детерминации. Т.о., взаимодействие сигналов FGF и Notch в передней части psm м. закладывать дефинитивные ростральные границы Hox. Задние трансформации, достигаются экспериментально у трансгенных мышей с помощью эктопической более передней экспрессии Нох генов (Kessel et al., 1990; Lufkin et al., 1992; McLain et al., 1992). Скорее всего Lfng вызывает задние трансформации и передний сдвиг экспрессии Hoxb6. Это указывает на то, что активность Lfng в psm необходима для предотвращения эктопической активации или распространения экспрессии гена Hox кпереди от своей обычной границы экспрессии. Т.о., формально циклический Lfng репрессирует транскрипцию Hox генов во время закладки дифинитивных передних границ экспрессии.
Всё это демонстрирует, что как снижение передачи сигналов Notch без обнаружения нарушений экспрессии циклических генов в psm таки и нарушение циклической активности Lfng затрагивают позиционную спецификацию мезодермальных производных вдоль переден-задней оси тела. Т.о., передача сигналов Notch и более вероятно их циклическая модуляция необходимы для спецификации качественных особенностей позвонков.
Сайт создан в системе
uCoz