The canonical Wnt pathway in early mammalian embryogenesis
and stem cell maintenance/differentiation
Jianbo Wang and Anthony Wynshaw-Boris Current Opinion in Genetics & Development 2004, 14:533–539
The Wnt signaling pathway plays essential roles in embryogenesis of higher eukaryotes from diploblastic, radially symmetrical cnidarians to mice and humans. Whereas studies in model organisms such as Drosophila and Xenopus continue to elucidate how the Wnt pathway is regulated,
studies of mouse mutants and cultured mammalian cells start to reveal how the Wnt pathway controls development and differentiation in mammals. Here we review some of the recent
progresses in our understanding of the Wnt pathway. We focus on how this pathway, through regulating transcription of its downstream target genes, specifies pattern formation during
mammalian embryogenesis and functions in the differentiation and maintenance of stem cells both in vivo and in vitro.
Wu, X. et al.
Rac1 activation controls nuclear localization of β -catenin during canonical Wnt signaling.
Cell 133, 340–353 (2008)
Исследователи установили, что малая GTPase Rac1 является критическим компонентом пути канонической передаси сигналов Wnt. Fanxin Long с коллегами показали, что в ответ на Wnt, Rac1 активирует Jun N-terminal kinase-2 (JNK2), которая фосфорилирует β-catenin и способствует его локализации в ядро.
Работая на стромальной клеточной линии (ST2), которая диффернецируется в остеобласты в ответ на WNT3a, авт. наблюдали. что WNT3a активирует Rac1. Экспрессия доминантно-негативной формы Rac1 или RNA interference-обусловленное истощение Rac1 ингибирует экспрессию WNT3a репортерного гена и предупреждает дифференцировку ST2. В отсутствие Rac1, эндогенный β-catenin не накапливается в ядре в ответ на передачу сигналов Wnt. Итак, Rac1 необходима для активности каноническго Wnt сигнального пути вов время дифференцировки остеобластов, но как Rac1 контролирует ядерную локализацию β-catenin?
Доминантно-негативная Rac1 ингибирует активацию JNK2 , а химическое ингибирование JNK2 предупреждает накопление в ядре β-catenin и снижает экспрессию репортерного гена в ответ на WNT3a. Используя биохимические методы, авт. показали. что β-catenin, Rac1 и JNK2 взаимодействуют в цитозоле ST2 клеток, Более того, в ответ на WNT3a, JNK2 фосфорилирует Ser191 и Ser605 в β-catenin;, фосфорилированный β-catenin затем мигрирует в ядро.
Каноническая передача сигналов Wnt вжна для роста конечностей. Чтобы определить физиологическое значение Rac1 для передачи сигналов Wnt, авт. генетически устраняли Rac1 из apical ectodermal ridge (AER) зачатков эмбриональных конечностей мышей. Эмбрионы, лишенные Rac1 в AER имели дефекты укорочения конечностей, которые были идентичны тем, что ранее наблюдались у β-catenin мутантов. Более того, используя репортерную линию мышей, которая была получена, чтобы отражать передачу сигналов β-catenin in vivo, группа показала, что передача сигналов β-catenin не индуцируется в AER в отсутствие Rac1. в
Наконец, эмбрионы, лишенные одной копии Rac1 и β-catenin имели идентичные дефекты с теми, что у β-catenin или Rac1-нокаутных эмбрионов. Сходные результаты были получены при удалении одной копии Rac1 и избыточной экспрессии Dickkopf-1 (Dkk1), который как известно, противодействует каноническому сигнальному Wnt пути. Эти находки указывают на то, что Rac1 генетически взаимодействует с β-catenin и Dkk1, чтобы контролировать рост конечностей.
Итак, эти данные предоставляют перые доказательства, что Rac1 контролирует канонический сигнальный Wnt путь и выявляют давно популярный механизм, который регулирует транслокацию в ядро β-catenin: в ответ на передачу сигналов Wnt Rac1 активирует JNK2, которая фосфорилирует β-catenin. Эта модификация заставляет его перемещаться в ядро. Играет ли Rac1 сходную роль в др. физиологических событиях, помимо развития эмбрионального AER конечностей, необходимо исследовать.
Li, F.-Q. et al. Chibby cooperates with 14-3-3 to regulate beta-catenin subcellular distribution and signaling activity. J. Cell Biol. 181, 1141–1154 (2008)
Wnt–β-catenin путь является жизненно важным для многочисленных клеточных событий во время эмбрионального развития, тканевого гомеостаза и туморогенеза. β-catenin проникает в ядро вследствие стимуляции Wnt и действует как транскрипционный ко-активатор благодаря связыванию T-cell factor/lymphoid enhancer factor (TCF/LEF). Импорт и экспорт в ядро β-catenin представляет собой критическую ступень в регуляции уровней β-catenin, компетентных обеспечивать передачу сигналов, т.к. белок осуществляет свою сигнальную активность только в ядре. Takemaru с коллегами открыли новый механизм, который контролирует динамику трафика β-catenin между ядром и цитоплазмой.
Группа Takemaru ранее описала β-catenin антагонист, Chibby (Cby), который эволюционно законсервирован от мухи до человека. Cby физически взаимодействует с β-catenin способом, который конкурирует с TCF/LEF, и т.о., репрессирует β-catenin-обеспечиваемую активацию генов. Снижение уровней Cby у эмбрионов мух с помощью RNA interference ведет к гиперактивации этого пути, что подчеркивает биологическую важность функции Cby.
Для дальнейшего выяснения молекулярной и клеточной функций Cby, Takemaru с коллегами изолировали Cby-связывающих партнеров. Используя affinity purification-mass spectrometry, они идентифицировали 14-3-3ε и 14-3-3ζ в качестве новых факторов, взаимодействующих с Cby. 14-3-3 белки, которые соединяются с множеством функционально разнообразных сигнальных белков, специфически распознают Ser20 на N-терминальном 14-3-3-связываюшем консенсусном мотиве у Cby. Это взаимодействие нуждается в фосфорилировании этого критического остатка Ser с помощью AKT/protein kinase B (PKB), т.к. kinase-dead AKT/PKB мутант почти полностью лишен способности связываться 14-3-3 белков с Cby. Дикого типа Cby или локализуется в цитоплазме или как в ядре, так и цитоплазме, тогда как Cby мутантны, дефектные по связыванию 14-3-3 преимущественно локализуются в ядре. Эти находки указывают на то, что такая ассоциация с 14-3-3 запускает секвестрацию Cby в цитоплазме.
Авт. затем показали, что Cby и 14-3-3 формируют комплекс из трех молекул с β-catenin в клетках тканевой культуры. Интересно, что Cby и 14-3-3 сотрудничают, чтобы обеспечить цитоплазматическую локализацию β-catenin и тем самым ингибировать передачу сигналов β-catenin. при замене на Ala Ser20 в Cby элиминируется связывание с 14-3-3 и частично нарушается способность Cby ингибировать передачу сигналов β-catenin.
Несколько линий доказательств указывают на то, что AKT/PKB играет позитивную роль в передаче сигналов Wnt-β-catenin. Это исследование предоставило доказательства, что доставленный в ядро AKT/PKB ингибирует, тогда как связанный с мембраной AKT/PKB стимулирует обусловленную β-catenin активацию транскрипции. Возможно, что мембранная и цитоплазматическая AKT/PKB преимущественно фосфорилирует и тем самым инактивирует glycogen synthase kinase-3, тогда как ядерная AKT/PKB фосфорилирует β-catenin и Cby. В свою очередь это облегчает связывание 14-3-3 и последующее ядерное исключение тройного комплекса. Следовательно, субклеточная компартментализация AKT/PKB по-разному влияет на передачу сигнлов β-catenin. Эта находка подчеркивает сложную природу взаимодействий между Wnt и AKT/PKB сигнальными путями.
Итак, полученные данные указывают на новую парадигму, согласно которой Cby действует совместно с 14-3-3 белками, чтобы облегчить экспорт в ядро β-catenin и тем самым противодействовать передаче сигналов β-catenin. Авт. полагают, что ингибирование передачи сигналов β-catenin с помощью Cby участвует, по крайней мере. в двух самостоятельных механизмах: в конкуренции TCF/LEF транскрипционных факторов за β-catenin в ядре и за облегчение экспорта в ядро β-catenin посредством взаимодействия его с 14-3-3 после своего фосфорилирования с помощью AKT/PKB.
Хотя и был открыт в качестве онкогена 22 года тому назад [1], но наше понимание Wnts и сигнальных событий с ними связанных получены в основном на основании генетических и онтогенетических исследований у таких модельных организмов как Drosophila и Xenopus. Идентифицированы многие ключевые компоненты и вычленен канонический путь: связывание Wnt со своим рецептором Frizzled (Fz) и ко-рецептором Arrow/Lrp5/6 (lipoprotein receptor related protein) увеличивает цитоплазматический уровень и накопление в ядре β-catenin. β-Catenin м. затем связывать транскрипционные факторы, такие как Tcf/Lef (T-cell transcription
factor/lymphocyte enhancer factor), чтобы активровать экспрессию генов мишеней [2-4]; см http://
www.stanford.edu/~rnusse/pathways/celldouble.html схематическую диаграмму).
Открыто несколько точек дивергенции от пути Wnt. Напр., Wnt5 м. служить в качестве сигнала наведния аксонов путем соединения с тирозин киназным рецептором у мух [5], так что лиганд полностью не связанный с Wnt м. активировать β-catenin обусловленную транскрипцию путем специфического связывания Fz4 рецептора у млекопитающих [6]. Тем не менее, ключевые цитоплазматические события канонического Wnt пути, т.e. Gsk3β; (glycogen synthase kinase 3β) обусловленная деградация β-catenin, селективная стабилизация β-catenin, ядерная локализация и последующая трансактивация, скорее всего происходят очень рано и приспособлены для регуляции процесса гаструляции, фундаментального онтогенетического события, которое создает зародышевые листки [7]. О неканонических Wnt путях с передачей сигналов Rho/Rac иd Ca2+ подробно говорится в обзоре Veeman et al. [8].
The canonical Wnt pathway
Центральным игроком в передаче сигналов Wnt является β-catenin.
Однако, β-catenin не является исключительным только для Wnt
пути, а выполняет и др. важную функцию в цитоплазме: формирование cadherins соединений [9]. В более специализированных клетках, таких как нейроны или клетках, проходящих митоз, β-catenin выполняет дополнительные критические функции [10-12]. Т.о., предпосылкой передачи сигналов Wnt является то, что в клетках, не подвергающихся воздействию Wnt, уровень и локализация β-catenin регулируются тонко для предупреждения активации транскрипции, тогда как в то же самое время обеспечивается поддержка др. клеточных функций. Не удивительно, что множественные механизмы используются для достижения этой цели.
Наиболее охарактеризован и понятен наиболее консервативный [7] Gsk3β-обусловленного фосфорилирования β-catenin и его последующая деградация с помощью протеосом, чтобы поддерживать уровнеь свободного β-catenin на низком уровне. Axin and APC (adenomatous poliposis coli) также необходимы для этого процесса, чтобы сформировать 'комплекс деструкции' с Gsk3β (reviewed in [2-4,13]). Однако, у эмбрионов Drosophila если уровень эндогенного β-catenin понизить с помощью двух гипоморфных аллелей, то потеря Gsk3β больше не вызывает повсеместной активации β-catenin и клетки восстанавливают свою способность отвечать на Wnt, указывая тем самым на то, что существует Gsk3β-независимая супрессия активности β-catenin. В такой ситуации, оставшаяся супрессия β-catenin приписывается Axin [14]. Механизм Axin обусловленной Gsk3β-независимой супрессии активности β-catenin неизвестен, хотя предполагается закрепляющая в цитоплазме активность Axin совместно с APC [13]. Однако, тот факт, что избыточно экспрессирующийся Axin сохраняет некоторую активность по супрессии передачи сигналов Wnt APC нулевых эмбрионов мух не совсем согласуется с этой гипотезой [15]. Cong and Varmus показали на культуре ткани, что экзогенный Axin снуёт между ядром и цитоплазмой и м. способствовать изменению локализации β-catenin из ядра в цитоплазму, указывая на ещё неизвестный механизм, с помощью которого Axin м. супрессировать активацию транскрипции [16].
Поиск партнеров, связывающих β-catenin, выявил третьий механизм, используемый клетками для супрессии активации генов-мишеней с помощью β-catenin в отсутствие стимуляции Wnt. Ядерный белок, Chibby, м. конкурировать с транскрипционными ко-факторами Tcf/Lef по связыванию β-catenin, чтобы репрессировать транскрипционную активацию. Анализ потери функции у эмбрионов мух подтвердил, что этот антагонист обязателен, даже если все др. механизмы ингибирования β-catenin всё ещё функционируют [17]. Т.о., множественные механизмы необходимы для супрессии активности β-catenin в отсутствие стимуляции Wnt. Активация передачи сигналов Wnt характеризуется накоплением β-catenin в ядре. Неясно, существует ли механизм, который способствует транспорту в ядро β-catenin после Wnt стимуляции. Однако, если Axin в самом деле активно сдвигает локализацию β-catenin в цитоплазму [16], то блокирование активности Axin м.б. достаточно для обратного накопления β-catenin в ядре. Более того, т.к. Axin необходим для функционирования комплекса деструкции, то манипуляции со стабильностью, локализацией или активностью Axin будут приводить к накоплению β-catenin в цитоплазме. В подтверждение этому было показано, что повторяющийся PPPSP мотив в цитоплазматическом хвосте ко-репрессора Arrow/Lrp м. фосфорилироваться после Wnt стимуляции, чтобы сформировать индуцибельный docking сайт для Axin [18]. Рекрутирование Axin в мембрану с помощью Arrow/Lrp м. инактивировать комплекс деструкции или путем деградации [13,14] или секвестрации Axin в клеточной мембране [15]. Используя экстракты из яиц лягушек, Lee et al. нашли, что концентрация Axin на несколько порядков величин ниже по сравнению с др. компонентами пути Wnt, включая APC и Gsk3β, которые также являются компонентами нескольких др. важных путей. Их математическая модель подтверждает, что с помощью манипуляций с Axin, передача сигналов Wnt м. оперировать с минимальными изменениями в уровнях APC и Gsk3β, вызывая тем самым меньше пертурбаций в др. путях [19].
Др. ключевой игрок в активации Wnt это белок Dishevelled (Dsh or Dvl), необходимый как для канонического, так и не-канонического Wnt пути [8,20]. Dsh/Dvl , как полагают, 'активируется' после Wnt стимуляции. Несмотря на тот факт, что выявлены многие партнеры по связыванию [20] и наблюдается повышенное фосфорилирование после Wnt стимуляции, механизм активации Dsh/Dvl всё ещё неясен. Исходя из их наблюдения, что Axin-GFP трансген обнаруживает Dsh/Dvl зависимую локализацию в мембране, Cliffe et al. предложили модель, согласно которой Dsh/Dvl постоянно переносит Axin-содержащий комплекс деструкции в плазматическую мембрану, где он м.
удерживаться с помощью Arrow/Lrp после Wnt стимуляции [15]. Более того, из-за того, что PDZ домен у Dsh/Dvl м. соединяться с цитоплазматической порцией рецептора Fz непосредственно [21], он м. также доставлять Axin в клеточную мембрану независимо от LRP5/6/Arrow [3].
В конечном итоге β-catenin накапливается в ядре, где он соединяется с промоторами своих ниже стоящих генов мишеней путем взаимодействия с транскрипционными факторами семейства Tcf/Lef. β-catenin м. смещать транскрипционные ко-репрессоры, такие как Groucho и histone deacetylase (HDAC), рекрутируя при этом histone acetyltransferase CBP/p300 и хроматин-ремоделирующий энзим Brg-1, чтобы обеспечить транскрипционную активацию [4,22,23]. недавно было показано, что два ядерных белка, Legless и Pygopus, необходимы для активации транскрипции β-catenin [2,4]. В отличие от CBP/p300 и Brahma, которые соединяются с C-терминальной областью β-catenin, Legless
соединяется с N-терминальными Armadillo повторами в β-catenin. Он затем рекрутирует Pygopus, который м. служить как трансактиватор. Будучи прикреплен непосредственно к Tcf, Pygopus активировать Wnt гены мишени в отсутствие β-catenin [24].
Хотя роль β-catenin в Wnt пути безусловно ядерная [25-27], Briata et al. выявили второй слой регуляции экспрессии Wnt генов мишеней, который, по-видимому, осуществляется в цитоплазме. Используя линию гипофизарных клеток, они показали, что воздействие Li или избыточная экспрессия активированного β-catenin м. специфически стабилизировать мРНК ряда известных транскрипционных мишеней Wnt, включая Pitx2, c-Jun, CyclinD1 и D2. Этот эффект обеспечивается путем усиления переноса ядро->цитоплазма HuR, мРНК стабилизирующего adenylate/uridylaterich
element (ARE) связывающего белка и одновременно переносом цитоплазма->ядро KSRP и TTP, мРНК дестабилизирующих ARE-связывающих белков. Более того белок Pitx2 сам по себе играет критическую роль в этом процессе. т.к. избыточная экспрессия дикого типа Pitx2 достаточна для стабилизации мРНК до уровня, сходного с таковым при воздействии Li, тогда как избыточная экспрессия мутантной формы Pitx2 м. доминантно дестабилизировать мРНК даже при воздействии Li [28]. Это демонстрирует существование нового механизма, при котором транскрипционные мишени Wnt/β-catenin м.б. активированы умноженным и скоординированным образом.
Wnt signaling in mammalian development:
axis formation and early patterning
хотя исследования на модельных организмах, таких как Drosophila
и Xenopus выявили существенную роль передачи сигналов Wnt в эмбриогенезе, не всегда возможно непосредственно экстраполировать функционирование передачи сигналов Wnt в контроле развития и дифференцировки у млекопитающих.
Имеется, по крайней мере, 19 Wnt лигандов, 10 Fz рецепторов и два ко-репрессора (Lrp5 и 6) у мышей (http://
www.stanford.edu/~rnusse/wntwindow.html), это указывает на значительное генетическое перекрывание затрудняет функциональную характеристику передачи сигналов Wnt. Это скорее всего происходит во время раннего эмбриогенеза (т.e. gastrulation), когда некоторые Wnts обнаруживают перекрывающиеся паттерны экспрессии [29]. В этой связи, удивительно, что мутации одиночных Wnt, Wnt3, нарушают гаструляцию путем блокирования первичной полоски и , следовательно, формирование мезодермы и дефинитивной энтодермы [30] (Рис. 1). Lrp5/6 двойные мутанты, также как и классический мутант mesd (mesoderm development) мышей, который делетирует ген, кодирующий Mesd, специфический хаперон для Lrp5 и 6, обладают очень сходными дефектами паттерна, как и Wnt3 мутанты [31,32]. Напротив, потеря функции β-catenin дает в результате дополнительные дефекты помимо неспособности образования первичной полоски [33,34,35]. В отличие от Wnt3, Lrp5/6 и mesd мутанты, у которых передняя спецификация нейроэктодермы с помощью AVE (anterior visceral endoderm) , по-видимому, происходит в отсутствие первичной полоски, β-catenin мутанты не способны ориентировать дистальную висцеральную энтодерму в переднюю сторону
(Рис. 1). Анализ микромассивов и in situ показал, что экспрессия Cripto, гена, кодирующего ко-рецептор для передачи сигналов Nodal, являющаяся существенной для передней транслокации дистальной висцеральной энтодермы, отсутствует в эпибласте β-catenin мутантов [35]. Дальнейшие эксперименты с β-catenin мутациями с избыточностью функции и подходы с репортерными генами указывают на то, что Cripto м.б. непосредственной мишенью комплекса Lef/Tcf/β-catenin [35]. Из-за важной роли Arrow/Lrp5/6 в передаче Wnt сигналов, как указывалось выше, фенотипические различия между β-catenin и Lrp5/6 или mesd мутантами указывают на Wnt-независимую транскрипционную активацию с помощью β-catenin в формировании переднего организатора у эмбрионов мышей [35]. необходимо отметить, что , по крайней мере, на ранних стадиях эмбриогенеза, когда проводился анализ, удаление β-catenin, по-видимому, не меняет межклеточной адгезии, преимущественно из-за компенсаторной замены plakoglobin [33,34].
Довольно интригующе, но согласующаяся с экспериментами на лягушках и рыбках данио, последующая индукция переднего мозга нуждается в ингибировании передачи сигналов Wnt [29,36-39]. Как AVE , так и аксиальная мезодерма, происходящие из передней части первичной полоски, экспрессируют Dickkopf1 (Dkk1), секретируемый антагонист Wnt, который соединяется с Lrp5/6. Анализ химер, однако, показал, что экспрессия Dkk1 в AVE не существенна, а Dkk1, секретируемый в AME, достаточен для формирования передней части головы [39]. Исследования на курах и Xenopus подтвердили, что ингибирование передачи сигналов Wnt является также критическим и для формирования сердца [29,38]). В согласии с этими исследованиями, эксперименты с химерами и условными делециями у мышей показали, что β-catenin, а , следовательно, и предача сигналов Wnt не требуют клеточно автономного проявления в клетках эпибласта, чтобы внести вклад в кардиальную мезодерму [33,34]. Однако, у эмбрионов мышей, несущих гипоморфные мутации Apc, передний мозг укорочен, но кардиальная мезодерма м.б. специфицирована, судя по экспрессии маркерного гена, хотя в аномальной дорсальной локализации [36]. У химерных эмбрионов дикого типа вне-эмбриональные ткани м. восстанавливать неправильно локализованную кардиальную мезодерму у Apc гипоморфных мутантов, чтобы сформировать нормальные сердца, но не укорочение переднего мозга [36]. Т.о., ингибирование Wnt inhibition при формировании сердца у млекопитающих нуждается в более осторожном обращении в будущем.
В противоположность нейроэктодерме и мезодерме, β-catenin необходим для спецификации энтодермы. В отсутствие β-catenin энтодермальные клетки меняют свою судьбу и формируют кардиальную мезодерму [34]. Как β-catenin
осуществляет выбор судьбы между энтодермой и мезодермой, неясно.
Wnt signaling in progenitor and stem
cells in vivo
Исследования формирования паттерна головного мозга у кур привели к модели, согласно которой повышенные уровни Wnt вдоль передне-задней оси наделяют нейральные клетки с первоначально ростральными характеристиками градиентно более каудальными характеристиками [40]. В соответствии с этой моделью продолжающееся ингибирование передачи сигналов Wnt у мышей необходимо для спецификации переднего мозга после инициальной индукции головы. Транскрипционный фактор Six3 действует на этой ступени, по крайней мере, частично посредством ингибирования транскрипции Wnt1, чтобы поддерживать качественные особенности клеток, предназначенных судьбой формировать передний мозг [41]. Роль Wnt в формировании каудальных частей головного мозга мышей не изучена. Эксперименты по генному таргетингу выявили только, что Wnt1 и Wnt3a участвуют в клеточной пролиферации в диэнцефалоне, среднем и заднем мозге [42]. Во время последующего развития передача сигналов Wnt/β-catenin необходима для всей ЦНС для увеличения популяции клеток предшественников с помощью одновременного стимулирования клеточной пролиферации и блокирования апоптоза и дифференцировки [43].
A schematic diagram of early mouse embryogenesis. Phenotypic characterization of several mouse mutants suggest that whereas Wnt3 activates bcatenin through co-receptors Lrp5 and 6 to promote primitive streak formation in the posterior part of embryo, b-catenin may function in a ligandindependent fashion to orient distal visceral endoderm anteriorly to form AVE. (See main text for detail.)
Как в точности β-catenin способствует клеточной пролиферации в нервной системе остается неизвестным. В гипофизарных нейронах и клетках кардиального нервного гребня β-catenin м. увеличивать транскрипцию Pitx2 путем соединения с и смещением HDAC1 от Lef1, связанного с Pitx2промотором. Затем β-catenin превращает Pitx2 из репрессора в активатор путем замещения HDAC1 и тем самым активирует Cyclin D2 для помощи пролиферации [22]. Важно отметить, что этот механизм является специфическим для типов клеток. В раннем нативном нервном гребне β-catenin не оказывает влияния на клеточную пролиферацию, а скорее индуцирует специфически судьбу сенсорных нервных клеток [44].
Исследования линий клеток colorectal рака и трансгенных мышей, которые эктопически экспрессируют Dkk1, также продемонстрировало существенную роль передачи сигналов Wnt в поддержании стволовых клеток кишечника, содействуя пролиферации и блокируя дифференцировку [45-47]. Это достигается, по крайней мере, частино посредством активации Tcf-4 гена-мишени c-Myc, который в свою очередь репрессирует транскрипцию p21CIP/WAF1 [45,46]. В культивируемых C2C12 мышечных клетках, β-catenin м. активировать транскрипцию c-Myc путем связывания Pitx2 aи E2F/p130 pocket белкового ингибирующего комплекса, чтобы отделить HDAC1 от промотора [23]. Способность β-catenin активировать Pitx2 обеспечиваемую транскрипцию G1 рост контролирующих генов cyclin D1 и D2 в клетках рака толстой кишки, м. указывать на дополнительный механизм для передачи сигналов Wnt для обеспечения пролиферации стволовых клеток [23].
Интересный поворот происходит во время формирования волосяных фолликулов, где функция β-catenin атипична в зачатках волосяных фолликулов, происходящих из разрастаний стволовых клеток эпителия кожи. В противоположность своей конвенционной роли в конвертировании Lef/Tcf из транскрипционных репрессоров в активаторы, β-catenin репрессирует транскрипцию E-cadherin, благодаря связыванию Lef1 [48]. Пониженные уровни E-cadherin и ослабление межклеточной адгезии скорее всего являются критическими для морфогенеза фолликулов [48].
Wnt signaling in stem cells in vitro
Роль передачи сигналов Wnt в поддержании и дифференцировке мультипотентных стволовых клеток выявлена на очищенных из костного мозга haematopoietic stem cells (HSCs) [49,50]. Усиление передачи сигналов Wnt посредством или ретровирусной инфекции стабилизированного β-catenin или воздействия очищенного белка Wnt3a, блокирует дифференцировку и способствует само-обновлению, а также способности восстанавливать клетки крови, когда трансплантируются летально облученным мышам. Существенная роль передачи сигналов Wnt в поддержании судьбы само-обновляющихся HSCs в дальнейшем подтверждена наблюдениями, что блокирование предачи сигналов Wnt с помощью избыточной экспрессии или Axin или секретируемой формы Fz cysteine-богатого домена, которая конкурирует с эндогенным Fz рецептором для связывания Wnt, ингибирует пролиферацию in vitro и способность HSCs восстанавливать клетки крови in vivo. Напротив, производные мышц CD45+ стволовые клетки - которые обладают потенциалом дифференцироваться в HSC или миогенные предшественники - в смешанной Wnt5a, 5b, 7a и 7b кондиционной среде м. специфически способствовать миогенной дифференцировке этих клеток [51]. Возможно, что передача сигналов Wnt м. действовать на разные транскрипционные мишени в разных контекстах. В мышечных стволовых клетках, промотор Pax7 содержит несколько Tcf/Lef сайтов связывания и предполагается, что является непосредственной мишенью для передачи сигналов Wnt для последующей детерминации в миогенный клон [51]. Напротив, гены, участвующие в само-обновлении HSC, такие как HoxB4 м Notch1, как было показано усиливают свою активность после активации передачи сигналов Wnt в HSCs [50]. Являются ли эти гены прямыми мишенями для Wnt сигналов, неизвестно.
Conclusions
Over the past two decades, there have been major
advances in our understanding of Wnt signaling during
mammalian embryogenesis, owing to the use of multiple
model organisms. In vitro studies in cell culture have shed
light on the transcriptional targets through which Wnt
signaling controls differentiation and stem-cell maintenance.
A central question one may then ask is: how does
the Wnt pathway regulate specific transcriptional targets
in different cells at different times to achieve distinct
outcomes? A better understanding of the integration
between Wnt and other signaling pathways and the
impact of epigenetic modifications on the transcription
of Wnt target genes may help answer this question in the
future.

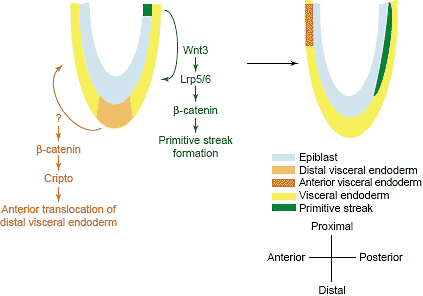 A schematic diagram of early mouse embryogenesis. Phenotypic characterization of several mouse mutants suggest that whereas Wnt3 activates bcatenin through co-receptors Lrp5 and 6 to promote primitive streak formation in the posterior part of embryo, b-catenin may function in a ligandindependent fashion to orient distal visceral endoderm anteriorly to form AVE. (See main text for detail.)
A schematic diagram of early mouse embryogenesis. Phenotypic characterization of several mouse mutants suggest that whereas Wnt3 activates bcatenin through co-receptors Lrp5 and 6 to promote primitive streak formation in the posterior part of embryo, b-catenin may function in a ligandindependent fashion to orient distal visceral endoderm anteriorly to form AVE. (See main text for detail.)