Простота небольших пресноводных полипов гидр с ее единственной осью и с почти безграничной способностью регенерации инспирировала теорию формирования биологического паттерна ([4]; см. также [3]). Ключевым предположением было то, что формирование паттерна м. происходить если и только если локальная сама себя усиливающая петля обратной связи связана с антагонистической реакцией на более длинных расстояниях (Рис. 1). Простая реализация такого взаимодействия заключается в аутокаталитическом активаторе, который продуцирует дально-действующий ингибитор, который, в свою очередь, противодействует само-активации. В такой интерпретации, гомогенное распределение является нестабильным. В возникающей области высокая концентрация активатора м. действовать как организатор (Рис. 4). Тем временем, некоторые такие пары активатор-ингибитор были обнаружены в разнообразных системах (Рис. 1). Паттерн м.б. ориентирован так за счет матерью обеспечиваемой асимметрии. Паттерны являются крепкими и в основном не зависят от инициальных условий.
Модели характеризовались высокой степенью саморегуляции, т.к. они наблюдались после экспериментальных вмешательств (Рис. 1c). Долгое время исследования по формированию осей у высших организмов редко соотносили с экспериментами по формированию паттерна у гидр, хотя Coelenterates, в целом, считались близкими к последнему общему предшественнику перед возникновением билатерального плана тела в ходе эволюции. Гидра, скорее всего, д. выявлять некое архитипическое формирование оси. Находка, что β-catenin и Wnt экспрессируются в органзаторе гидр
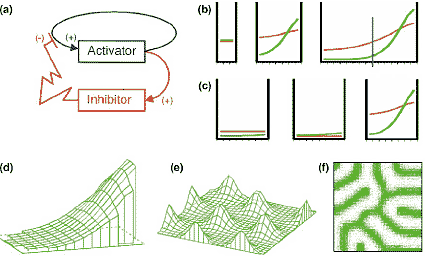 Primary pattern formation. (a) Reaction scheme: it was predicted that a short-ranging substance called the activator has a positive feedback on its own
production. This self-enhancement is antagonized, e.g. by a long-ranging inhibitor [3,4]. The Nodal/Lefty2 [40] is an interaction is of this type.
As frequently observed [41], the organizing region is also the source of the antagonist. (b) In growing fields, a single maximum arises at a marginal position whenever the field size has obtained a critical extension. Although the same rules go for all cells, graded distributions emerge that are convenient to supply positional information. (c) Pattern regulation: after removal of the organizing region (cut at the blue line in [b]), the remnant inhibitor decays and, due to autocatalysis, the pattern regenerates. (d) In a two-dimensional field, the gradient orients along the longest extension. (e) If the range of the inhibitor is smaller than the size of the field, periodic patterns are formed. The trichomes on leaves (see Pesch and HuЁ lskamp, this issue),feather patterns [42], spacing of heterocysts [43], neural precursor and sensory bristle patterns in Drosophila [44] are examples. If the autocatalysis saturates, stripe-like patterns are formed [29,45]. To obtain a single stripe-like organizing region as required for the DV organization, auxiliary
reactions are required (see Figure 4). For animated simulations see http://www.eb.tuebingen.mpg.de/meinhardt.
Primary pattern formation. (a) Reaction scheme: it was predicted that a short-ranging substance called the activator has a positive feedback on its own
production. This self-enhancement is antagonized, e.g. by a long-ranging inhibitor [3,4]. The Nodal/Lefty2 [40] is an interaction is of this type.
As frequently observed [41], the organizing region is also the source of the antagonist. (b) In growing fields, a single maximum arises at a marginal position whenever the field size has obtained a critical extension. Although the same rules go for all cells, graded distributions emerge that are convenient to supply positional information. (c) Pattern regulation: after removal of the organizing region (cut at the blue line in [b]), the remnant inhibitor decays and, due to autocatalysis, the pattern regenerates. (d) In a two-dimensional field, the gradient orients along the longest extension. (e) If the range of the inhibitor is smaller than the size of the field, periodic patterns are formed. The trichomes on leaves (see Pesch and HuЁ lskamp, this issue),feather patterns [42], spacing of heterocysts [43], neural precursor and sensory bristle patterns in Drosophila [44] are examples. If the autocatalysis saturates, stripe-like patterns are formed [29,45]. To obtain a single stripe-like organizing region as required for the DV organization, auxiliary
reactions are required (see Figure 4). For animated simulations see http://www.eb.tuebingen.mpg.de/meinhardt.
[5] (Рис. 2) стала очень интересной, т.к. тот же самый путь играет также критическую роль в формировании организатора у позвоночных [6]. Регуляторное поведение экспрессии β-catenin/Wnt близко к тому, что предсказывается для активатора в нашей базовой модели формирования паттерна. Для
Drosophila имеются доказательства позитивной петли на пути β-catenin/Wnt [7]. Эксперименты на гидрах показали, что β-catenin имеет более широкое распространение, чем Wnt (Рис. 2a),подтверждая, что β-catenin и Wnt не являются компонентами простой петли. Широкое распределение β-catenin, по-видимому, является предварительным условием для более узкого пика Wnt, указывая на более косвенный аутокатализ (Рис. 2h). В формировании края крыла
Drosophila секретируемый
wingless выполняет ингибирующую функцию по отношению к самому
wingless [8]. Возможно, что секретируемый Wnt играет аналогичную ингибирующую роль и у гидр.
Большинство морфогенетических полей формируются на обоих терминальных концах. Это имеет место и в случае гидр. Более детальная модель формирования паттерна гидры [9] постулирует, что донная часть гидры функционирует в качестве вторичного организатора, антипода области первичного организатора. Экспрессия гомолога транскрипционного фактора Nkx2.5 в подошве гидры обнаруживает паттерн регуляции, который и ожидается для подошвенного активатора [10]. Установлено, что этот ген является авторегуляторным [11]. Как и ожидалось согласно теории эта автоактивация оказалась не линейной, т.к. используются димеры. Моделирование показало, что авторегуляторный эффект подошвенного активатора м. распространяться. Будучи транскрипционным фактором, Nkx2.5 определенно м. не диффундировать. Однако, др. игрок в этом аутокатализе является диффундирующим пептидом, pedibin, который, по-видимому, распространяет аутокаталитическую активацию. Молекулярная основа, которая составляет эту петлю на базальном конце, все ещё неизвестна. Т.к. это имеет место и в случае системы β-catenin/Wnt, упомянутой выше, то еще не найдено дальнодействующего ингибирования.
Hydra and the patterning of higher
organisms
Сравнение генов, участвующих в становлении плана тела у гидр и высших организмов выявили неожиданную связь родоначальной и современной
anteroposterior (AP) оси. Гены, которые экспрессируются в стволе тела гидры имеют своих аналогов в головах высших организмов. Это указывает на то, что все тело радиально симметричного родоначальника задейстовано в головном мозге и сердце высших организмов, тогда как туловище является более поздним приобретением в ходе эволюции [12]. Подошва гидры соответствует наиболее передним структурам на что указывает экспрессия
Nkx2.5 [10]. Напротив, экспрессия Wnt и Brachyury вокруг gastric отверстия [5,13] указывает на то, что т.наз. голова гидры соответствует наиболее задним структурам, бластопору. У гидры Goosecoid и
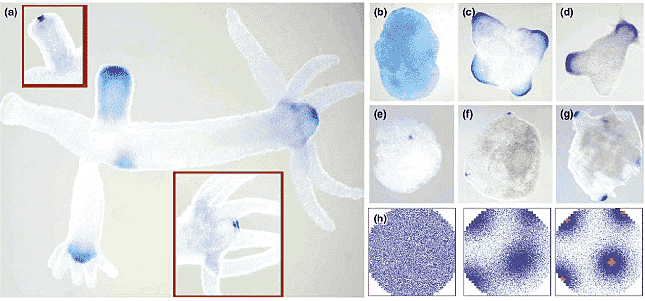 Organizer formation in hydra. (a) Hy-b-catenin and Hy-Tcf are expressed in the hypostome [5], the structure known from classical experiments to have organizer activity [1]. In the bud, a sharpening occurs in the course of time. Hy-Wnt expression (inserts) is more sharply confined to the oral opening than Hy-b-catenin. These maxima have many properties as expected from our model. (b-d) During pattern formation in re-aggregated cells, Hy-Tcf (and Hy-b-catenin) appears first uniformly distributed and become subsequently more restricted to regions that form eventually the heads. (e-g) In contrast, Hy-Wnt appears directly in sharp spots that form the future oral opening [5]. (h) The nested pattern formation can be accounted for by the assumption that both Hy-b-catenin/Hy-Tcf and Hy-Wnt are pattern forming systems and that a high Hy-b-catenin concentration is the precondition to trigger Hy-Wnt (red), which becomes more localized than the initiating Hy-b-catenin/Hy-Tcf pattern. (Photographs courtesy of B Hobmayer, T Holstein and colleagues; see [5].)
Organizer formation in hydra. (a) Hy-b-catenin and Hy-Tcf are expressed in the hypostome [5], the structure known from classical experiments to have organizer activity [1]. In the bud, a sharpening occurs in the course of time. Hy-Wnt expression (inserts) is more sharply confined to the oral opening than Hy-b-catenin. These maxima have many properties as expected from our model. (b-d) During pattern formation in re-aggregated cells, Hy-Tcf (and Hy-b-catenin) appears first uniformly distributed and become subsequently more restricted to regions that form eventually the heads. (e-g) In contrast, Hy-Wnt appears directly in sharp spots that form the future oral opening [5]. (h) The nested pattern formation can be accounted for by the assumption that both Hy-b-catenin/Hy-Tcf and Hy-Wnt are pattern forming systems and that a high Hy-b-catenin concentration is the precondition to trigger Hy-Wnt (red), which becomes more localized than the initiating Hy-b-catenin/Hy-Tcf pattern. (Photographs courtesy of B Hobmayer, T Holstein and colleagues; see [5].)
Brachyury экспрессируются по соседству с кольцами вблизи отверстия [13,14]. У позвоночных Goosecoid участвует в формировании паттерна головы [15], a Brachyury в формировании хвоста [16]. Это подтверждает, что туловище еще не присутствовало у родоначальных организмов, оно дифференцировалось позднее рядом с задним концом. Эта вставка туловища рекапитулируется во время онтогенетического развития. Рано в развитии позвоночных и Goosecoid и Brachyury экспрессируются в перекрывающихся областях [17], но оказываются разделенными в результате последовательной интеркаляции туловища. Сходным образом, у насекомых с коротким зародышевым щитком и у annelids голова формируется первой, тогда как сегментированное туловище добавляется позднее. Т.о. AP ось высших организмов состоит из двух частей и моделирование обеих частей нуждается в очень разных механизмах.
Родоначальная передняя часть (головной мозг и сердце) находятся под контролем градиента, который имеет свою наивысшую точку в бластопоре,
(т.e. в маргинальной зоне у
Xenopus или зародышевом кольце у рыб). У гидр сигнальные молекулы принадлежат семейству Wnt [18,19]. Это обусловливает заднюю трансформацию переднего default состояния (Рис. 3c и Chitnis, this issue). Напротив, формирование паттерна туловища м. объяснить, исходя из предположения, что имеет место осцилляция на заднем конце и что каждый полный цикл ведет к инсерции одной новой пары полу-сомитов (Figure 3f,g). Далее было предположено, что в купе с этой осцилляцией новые AP-специфицирующие гены (теперь это Нох гены) становятся активированными так, что генерируется не только периодический паттерн, но и метамерные элементы (сегменты или сомиты), получающие разные качественные AP особенности. Др. словами, осцилляция ведет к своего рода подсчету на генном уровне (Рис. 3g). Более 16 лет прошло с момента теоретического предсказания этого механизма [3] и экспериментального подтверждения [20,21].
A model for the generation of a near-
Cartesian coordinate system in vertebrates
Критической ступенью в развитии билатерального плана тела является установление вторичной оси, ортогонально по отношению к родоначальной AP оси. Все высшие организмы используют одну и ту же молекулярную систему спецификации (DV) оси [22]:
Chordin и BMPs (short gastrulation и dpp у
Drosophila). Обе молекулы уже присутствуют у гидр:
Chordin в головной области (T Holstein, F Rentsch, B
Hobmayer, personal communication), а BMP5/8 в области почек и в peduncle [23] (т.e. противоположно к области организатора). Это указывает на то, что система, которая обеспечивает формирование паттерна AP и DV осей у высших организмов уже присутствует у радиально симметричных родоначальников, хотя в общем-то в основном параллельно с др. Т.о., формирование вторичной оси у bilateria в основном не базируется на изобретении новой системы, а на взаимной переориентации
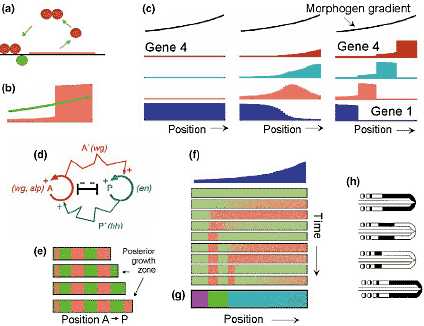 Models for gene activation, segmentation and somitogenesis. (a,b) A gene with an autocatalytic feedback on its own activation can form a switch ([46] and Slepchenko and Terasaki, this issue). Activation occurs whenever a signal (green) is above a certain threshold. Labial, Deformed and Twist are examples for genes with positive autoregulation [47-49]. (c) Activation of several genes by a single gradient was proposed to proceed by a stepwise irreversible promotion towards more posterior or more distal specifications [50], a dynamic observed, for instance, in hindbrain patterning [51,52]. (d) Prediction for segmentation: two or more feedback loops activate each other on long range but exclude each other locally [3,53]. The observed engrailed-wingless network turned out to be compatible with this prediction. (e) Example of a simulation: during outgrowth, the most posterior region alternates between two states, as observed for hairy in spiders [54]. (f) Somite formation was predicted to depend on an oscillation [3]. Each full cycle adds one pair anterior and posterior half-somitic specification (green/red). Signals move from posterior to anterior and come to rest at the position where the next half-somite is to be formed - that is, at a low level of a gradient (blue, now identified as FGF [55]) where oscillations are no longer possible. (g) Connected with this oscillation is a sequential activation of new AP-specifying genes (now HOX-genes), a prediction that is now also confirmed [21,55]. The periodic pattern and the AP-specifying genes are, therefore, precisely in register. (h) Observed c-hairy oscillation [20,56]: activity moves towards anterior and contributes to next posterior half somite to be formed (for modelling the oscillation, see Giudicelli and Lewis, this issue).
Models for gene activation, segmentation and somitogenesis. (a,b) A gene with an autocatalytic feedback on its own activation can form a switch ([46] and Slepchenko and Terasaki, this issue). Activation occurs whenever a signal (green) is above a certain threshold. Labial, Deformed and Twist are examples for genes with positive autoregulation [47-49]. (c) Activation of several genes by a single gradient was proposed to proceed by a stepwise irreversible promotion towards more posterior or more distal specifications [50], a dynamic observed, for instance, in hindbrain patterning [51,52]. (d) Prediction for segmentation: two or more feedback loops activate each other on long range but exclude each other locally [3,53]. The observed engrailed-wingless network turned out to be compatible with this prediction. (e) Example of a simulation: during outgrowth, the most posterior region alternates between two states, as observed for hairy in spiders [54]. (f) Somite formation was predicted to depend on an oscillation [3]. Each full cycle adds one pair anterior and posterior half-somitic specification (green/red). Signals move from posterior to anterior and come to rest at the position where the next half-somite is to be formed - that is, at a low level of a gradient (blue, now identified as FGF [55]) where oscillations are no longer possible. (g) Connected with this oscillation is a sequential activation of new AP-specifying genes (now HOX-genes), a prediction that is now also confirmed [21,55]. The periodic pattern and the AP-specifying genes are, therefore, precisely in register. (h) Observed c-hairy oscillation [20,56]: activity moves towards anterior and contributes to next posterior half somite to be formed (for modelling the oscillation, see Giudicelli and Lewis, this issue).
двух существующих систем формирования паттерна. Как это достигается?
У гидр gastric отверстие, бластопор, очень мало. Соотв. структура у позвоночных (напр., маргинальная зона у
Xenopus или зародышевое кольцо у рыб) является крупным кольцом. Тем не менее, у позвоночных родоначальная часть AP оси (головной мозг и сердце) организуется из (крупного) кольцеобразного бластопора, преимущественно с участием Wnt [18,19], полностью аналогично ситуации у гидр. Это не требует присутствия дорсального организатора типа Шпемановского [24]. Расширение небольшого заплатко-подобного организующего бластопора имеет важные последствия. consequence. Вторичное образование организатора Шпемановского типа на этом крупном кольце гарантирует вне-осевое положение, которое служит первой ступенью по направлению к созданию новой оси, перпендикулярной к первичной AP оси.
Однако, вне оси точко-подобный организатор недостаточен для DV-организации увеличвающегося в длину организма. Организующая область для DV оси д. распространяться на всю AP ось только с небольшим DV расширением. Т.о., такой организатор срединной линии д. иметь геометрию узкой полоски. Образование одиночной, длинной, но узкой исходной области нуждается в специфическом нетривиальном механизме [25,26]. Если некоторая латеральная ингибиция участвует в этом, то как срединная линия защищена от дизинтеграции на индивидуальных участках?
Моделирование показало, что одиночная узкая полоска м.б. генерирована, если небольшая точко-образная организующая область действует на систему формирования полоски таким образом, что м. возникнуть только одна полоска, будущая срединная линия. Разные стратегии используются в разных филетических группах [26] (Рис. 4). У позвоночных дорсальный организатор инициируется и удлиняется по срединной линии в переднем и заднем направлении. Раннее перемещение мезодермальных клеток происходит от бластопора кпереди. Клетки, которые оказались включенными в организатор формируют прехордальную пластинку и тем самым переднюю часть срединной линии. Это необходимо для формирования DV паттерна переднего- и среднего мозга [27].
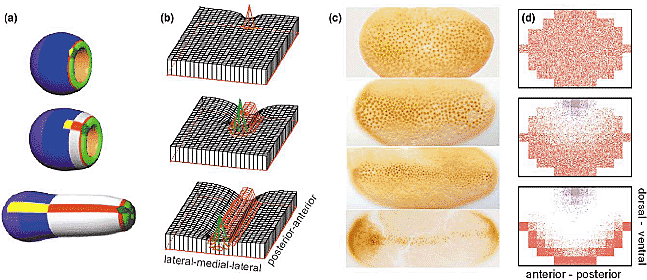 Different strategies for midline formation in vertebrates and insects. The organizing region for the DV organization has to extend over the whole AP axis
but must have only a small extension along the DV axis. Simple pattern-forming mechanisms leads either to patch-like or to multiple stripes-like distribution (see Figure 1d,f). Different phyla found different mechanisms to generate a single straight midline organizer. (a) In vertebrates, a Spemann-type organizer (dark green) is formed on the blastopore (light green) with a spot-like geometry. Midline formation occurs by elongation towards anterior (prechordal plate, yellow) and towards posterior (red). (b) Simulation: a spot-like organizer (node) triggers a stripe-forming system, which, in turn, repels the organizer and causes therewith its shift. In the course of time, the midline is left behind the moving node like a vapour trail behind a highflying airplane [25,26]. (c) In insects, the midline has from early stages onwards the full AP extension but sharpens along the DV axis (see [28] for review and Eldar, Shilo and Barkai, this issue); shown is the Dorsal pattern of the short germ insect Tribolium [57]. (d) Simulation: a dorsal patch-like organizer repels the a stripe-forming system. The single midline occurs ventrally [26,29]. Thus, the central nervous system forms in vertebrates on the same side as the organizer (dorsal), in insects on the opposite side (ventral). This may be the reason for the well-known DV-VD conversion between vertebrates and insects [30,31]. (Figure 4c courtesy of Siegfried Roth; see [57].)
Different strategies for midline formation in vertebrates and insects. The organizing region for the DV organization has to extend over the whole AP axis
but must have only a small extension along the DV axis. Simple pattern-forming mechanisms leads either to patch-like or to multiple stripes-like distribution (see Figure 1d,f). Different phyla found different mechanisms to generate a single straight midline organizer. (a) In vertebrates, a Spemann-type organizer (dark green) is formed on the blastopore (light green) with a spot-like geometry. Midline formation occurs by elongation towards anterior (prechordal plate, yellow) and towards posterior (red). (b) Simulation: a spot-like organizer (node) triggers a stripe-forming system, which, in turn, repels the organizer and causes therewith its shift. In the course of time, the midline is left behind the moving node like a vapour trail behind a highflying airplane [25,26]. (c) In insects, the midline has from early stages onwards the full AP extension but sharpens along the DV axis (see [28] for review and Eldar, Shilo and Barkai, this issue); shown is the Dorsal pattern of the short germ insect Tribolium [57]. (d) Simulation: a dorsal patch-like organizer repels the a stripe-forming system. The single midline occurs ventrally [26,29]. Thus, the central nervous system forms in vertebrates on the same side as the organizer (dorsal), in insects on the opposite side (ventral). This may be the reason for the well-known DV-VD conversion between vertebrates and insects [30,31]. (Figure 4c courtesy of Siegfried Roth; see [57].)
Задняя часть срединной линии инициируется и удлиняется организатором. который локализован в бластопоре (напр., узелке Гензена). Перемещающийся узел оставляет за собой срединную линию, как самолет оставляет за собой шлейф пара на высокой высоте. Это ведет к усадке кольца в пользу удлиняющейся аксиальной структуры - т.е., к превращению кольца бластопора, перпендикулярного к AP оси, в палочку, параллельную AP оси (Рис. 4a). Уменьшившееся кольцо оказывается самой задней структурой во время интеркаляции туловища.
У насекомых ситуация совершенно иная. Дорсальный организатор репрессирует образование срединной линии, позволяя образоваться срединной линии только на вентральной стороне [28,29]. С раннего развития срединная линия у насекомых полностью покрывает AP удлинение, но со временем она ограничивает DV ось (Рис. 4; см. Eldar, Shilo and Barkai, this issue). В то время как у позвоночных срединная линия и ЦНС формируются на одной и той же стороне организатора (дорсальной), у
Drosophila дорсальный организатор ограничивает формирование срединной линии противоположной (вентральной) стороной. Это м. дать новое объяснение хорошо известной конверсии DV-VD, отличающей насекомых и позвоночных [30,31].
Некоторые семейства генов у кораллов более сходны с таковыми у позвоночных, чем у насекомых или нематод [32]. Раннее разделение на стадии, когда обе оси были всё ещё параллельны др. др., происходило независимо для обеих осей без urbilaterian в качестве эволюционного промежуточного звена и м. внести неожиданные различия у сегодняшних высших организмов. Возможно, что инновации разных способов образования срединной линии были точкой невозвратности разделения разных phyla. В противоположность концепции urbilaterian, эти модели указывают на то, что вторичная ось существовала до возникновения билатеральности и что разные способы использовались для преобразования двух исходных параллельных осей и что одна из этих организующих областей получила геометрию полоски.
A possible link between the single axis of
hydra and appendages of higher organisms
Наиболее удивительным свойством является использование одних и тех же сигнальных систем для очень разных ступеней в развитии. Напр., β-catenin/wnt используется не только для генерации первичной оси тела, но и для инициации придатков у насекомых и позвоночных, зачатков перьев
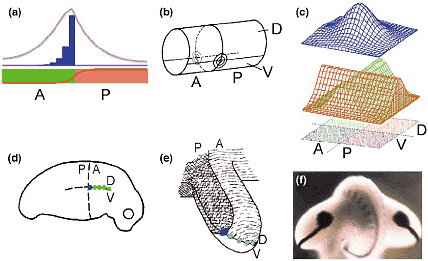 Boundary model for the organization of sub-structures such as legs and wings [58,59]. (a) By cooperation of differently determined cells (e.g. A and P;
see Figure 3), the border separating the two cell types becomes the source region for a morphogen. In this example a factor (brown) produced in
the P region is required in the adjacent A cells to produce the final morphogen. Its production is restricted to a narrow zone (blue) next to the
border and obtains (e.g. by diffusion) its final distribution (pink). Although the distribution is essentially symmetric to the border, both the A and the P region may respond differently. The hedgehog-dependent decapentaplegic production in the anterior compartment of the Drosophila wing disk is an example (see [60] for review). (b) Simplifying an embryo as a cylinder, the specification of a limb field requires the intersection of two borders, one along the AP and one along the DV axis (see Figure 4). Two signalling centres appear: one on the left and one on the right side. Both have opposite handedness. (c) By cooperation of the two signalling systems (red and green) produced along the two intersecting borders, a cone-shaped morphogen distribution (blue) is generated that is appropriate to organize the proximodistal axis. For insect legs it was predicted that three differently determined cell types have to cooperate to generate the positional information for distal outgrowth, anterior-dorsal, anterior-ventral and posterior [58], a prediction that turned out to be correct (see [61] for review). (d,e) For vertebrate limbs, it was proposed that only the anterior part is competent to respond to the morphogen [59]. The digits emerge along the DV border (large green dots), the type of the digits depends on the distance to the AP border. (f) View on a chick embryo from the tail onto the two wing buds [62]. The thick black lines result from a staining of FGF8 that is produced at the DV (Wnt7-a/En1) border (see [63,64] for review). The dark round spots mark high concentrations of Sonic hedgehog that is, according to the model, produced at an AP and DV border (blue spot in [d,e]). (Figure 5f kindly supplied by U Grieshammer [62].) The model found support from molecular genetic investigation [63] and accounts for many transplantation experiments performed in pre-molecular times [3,59].
Boundary model for the organization of sub-structures such as legs and wings [58,59]. (a) By cooperation of differently determined cells (e.g. A and P;
see Figure 3), the border separating the two cell types becomes the source region for a morphogen. In this example a factor (brown) produced in
the P region is required in the adjacent A cells to produce the final morphogen. Its production is restricted to a narrow zone (blue) next to the
border and obtains (e.g. by diffusion) its final distribution (pink). Although the distribution is essentially symmetric to the border, both the A and the P region may respond differently. The hedgehog-dependent decapentaplegic production in the anterior compartment of the Drosophila wing disk is an example (see [60] for review). (b) Simplifying an embryo as a cylinder, the specification of a limb field requires the intersection of two borders, one along the AP and one along the DV axis (see Figure 4). Two signalling centres appear: one on the left and one on the right side. Both have opposite handedness. (c) By cooperation of the two signalling systems (red and green) produced along the two intersecting borders, a cone-shaped morphogen distribution (blue) is generated that is appropriate to organize the proximodistal axis. For insect legs it was predicted that three differently determined cell types have to cooperate to generate the positional information for distal outgrowth, anterior-dorsal, anterior-ventral and posterior [58], a prediction that turned out to be correct (see [61] for review). (d,e) For vertebrate limbs, it was proposed that only the anterior part is competent to respond to the morphogen [59]. The digits emerge along the DV border (large green dots), the type of the digits depends on the distance to the AP border. (f) View on a chick embryo from the tail onto the two wing buds [62]. The thick black lines result from a staining of FGF8 that is produced at the DV (Wnt7-a/En1) border (see [63,64] for review). The dark round spots mark high concentrations of Sonic hedgehog that is, according to the model, produced at an AP and DV border (blue spot in [d,e]). (Figure 5f kindly supplied by U Grieshammer [62].) The model found support from molecular genetic investigation [63] and accounts for many transplantation experiments performed in pre-molecular times [3,59].
зубов [33-36], хотя последний общий предшественник наверняка не имел этих структур. Обычно такое повторное использование одной и той же сигнальной системы интерпретируется как привлечение доступной системы для новых целей. Формирование паттерна гидр и особенно их почкование (Рис. 2a) указывает, однако, на то, что не только сигнальная система
per se, но и полностью морфологический процесс повторно используется в модифицированной форме. Во время почкования возникает высокая локальная экспрессия β-catenin и Wnt, которая инициирует выросты для бесполого размножения (Рис. 2). Возможной ступенью в направлении развития придатков м.б. репрограммирование этих выростов в направлении не отсоединяющихся придатков. Minelli [37] уже предполагал, что конечности м. рассматриваться как модифицированные оси тела и он подчеркивал, что важной ступенью перехода от бесполого почкования к формированию придатков является замещение энтодермы почки мезодермой, т.к придаток не должен иметь ответвления от кишечной системы.
Инициация конечностей в качестве специфической дифференцировки стенок считается ключевой ступенью, благодаря которой конечности возникают в определенном положении с правильной полярностью, ориентацией и лево- право-ручностью (Рис. 5). Образование др. важных структур, таких как язык, базируется на локальном латеральном выросте энтодермы [38]. Является этот процесс раннего эктодермального и энтодермального ответвления в самом деле следствием бесполого почкования у родоначальных организмов покажет будущее. Напр., FGF играет главную роль в выделении легких, но мы пока не знаем, экспрессируется ли FGF у гидр. Подошва гидры сама по себе является примером иного использования части родоначального организма. Подошву м. рассматривать как родоначальное сердце [39]: она находится под контролем того же самого гена NKx2.5 [10], она инициируется в той же самой (очень передней) позиции и обладает активностью насоса, а частота её контракций м. зависеть от пептида, который также изменяет скорость сердцебиений у млекопитающих. Т.о., элементарные ступени формирования паттерна уже предсуществуют у радиально симметричных родоначальников и, по-видимому. используются в качестве строительных блоков для формирования
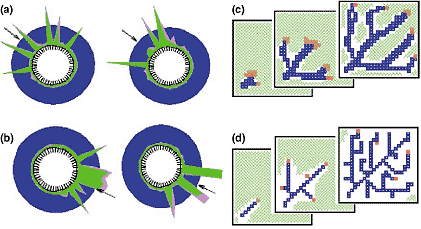 Model for the chemotactic orientation of cells and the generation of net-like structures. Some cells and growth cones are able to detect minute concentration differences to orient their movement (see [65,66] for review). (a) Model [67]: isolated signals (green) for filopods are generated by a (saturating) self-enhancing reaction and an inhibition that covers the whole cell. They appear preferentially at the side where the guiding signal (blue) is slightly higher. A second antagonistic reaction (pink) quenches the signal. Thus, signals for filopods appear and disappear permanently at the cell surface, monitoring the guiding signal. (b) Changes in the direction of the guiding signal are detected and new internal signals emerge at theappropriate side, although the guiding asymmetries are minute. If no guiding signal is available, protrusions continue to pop up, although at random positions, as observed. (c,d) Branching filaments can be generated if a local signal (red) causes filament elongation at the tip, either by cell proliferation, differentiation or single cell extension [3,46]. Elongation occurs towards a guiding factor (green) that is produced either ubiquitously or locally by a target region. It is removed by the filaments (blue). The filaments appear as a trace behind the moving signal. Branch formation either occurs by bifurcation (c) or by lateral branching (d) - that is, either the signal splits at the tip or a new signal appear along the filaments. The trachea of insects are an example [68,69] and most of the postulated components have observed counterparts.
Model for the chemotactic orientation of cells and the generation of net-like structures. Some cells and growth cones are able to detect minute concentration differences to orient their movement (see [65,66] for review). (a) Model [67]: isolated signals (green) for filopods are generated by a (saturating) self-enhancing reaction and an inhibition that covers the whole cell. They appear preferentially at the side where the guiding signal (blue) is slightly higher. A second antagonistic reaction (pink) quenches the signal. Thus, signals for filopods appear and disappear permanently at the cell surface, monitoring the guiding signal. (b) Changes in the direction of the guiding signal are detected and new internal signals emerge at theappropriate side, although the guiding asymmetries are minute. If no guiding signal is available, protrusions continue to pop up, although at random positions, as observed. (c,d) Branching filaments can be generated if a local signal (red) causes filament elongation at the tip, either by cell proliferation, differentiation or single cell extension [3,46]. Elongation occurs towards a guiding factor (green) that is produced either ubiquitously or locally by a target region. It is removed by the filaments (blue). The filaments appear as a trace behind the moving signal. Branch formation either occurs by bifurcation (c) or by lateral branching (d) - that is, either the signal splits at the tip or a new signal appear along the filaments. The trachea of insects are an example [68,69] and most of the postulated components have observed counterparts.
более сложных паттернов, наблюдаемых у высших организмов. Локальное само-усиление и дально действующее ингибирование играют ведущую роль.
Conclusions
Mathematically based modelling combined with computer
simulations is an efficient tool to find an inroad into the
complex web of interactions on which development is
based. The models reveal minimum interactions required
to accomplish particular steps. Hydra was originally chosen
as a model system to get an inroad into biological
pattern formation. Self-enhancement and long-range
inhibition was proposed to be the driving force in organizer
formation. More recent modelling suggests that this
radially symmetric animal also could be a key to understand
the establishment of the second axis and the
evolution of bilateral body plan. The body pattern of
the radially-symmetric animals evolved into the head
pattern of higher organisms. The long-extended but
narrow organizer required to pattern the dorsoventral
axis cannot be generated in a single step but requires
the cooperation of at least two patterning system. Different
phyla found different modes of this cooperation.
Whereas in vertebrates a dorsal organizer initiates and
elongates the midline at the dorsal side, in insects a dorsal
organizer repels the midline, which appears, therefore,
ventrally. The different modes of midline formation are
proposed to be decisive for the separation of phyla during
evolution.
Сайт создан в системе
uCoz