Complex redundancy to build a simple epidermal
permeability barrier
Julie Segre Current Opinion in Cell Biology 2003, 15:778–784
To survive the transition from an aqueous in utero to a terrestrial
ex utero environment, mice and humans must construct an
epidermal permeability barrier in utero. Terminally differentiated
epidermal cells, lipids and tight junctions are all essential to
achieve this barrier. Recent analyses of mouse mutants with
defects in structural components of the terminally differentiated
epidermal cell, catalyzing enzymes, lipid processing,
transcriptional regulators and the intercellular junctions have
highlighted their essential function in establishing the epidermal
permeability barrier. Particularly interesting examples include
modulation of the expression of transglutaminase 1 enzyme, the
transcription factor Klf4 and the claudin tight junction proteins.
However, careful analysis of the various mutant phenotypes
during embryonic development, as neonates and either as adults
or transplanted skin, has revealed much more about the
redundancy and compensatory mechanisms of the system.
Molecular analysis of the various mouse mutants has
demonstrated common pathways to compensate for loss of the
epidermal barrier.
Табл.1 Mouse mutants with defects in epidermal barrier permeability.
27. Kalinin AE, Kajava AV, Steinert PM: Epithelial barrier function:
assembly and structural features of the cornified cell envelope.
Bioessays 2002, 24:789-800.
Переход от водного in utero к земному образу жизни ex utero нуждается в полностью компетентном эпидермальном барьере проницаемости. Расположенный между телом и внешней средой эпидермис предупреждает как потерю жидкости, так и поступление токсических веществ. Незрелая кожа, которая развивает свои основные необходимые функции только на 34 неделе развития у людей, подвергает ребёнка значительному риску, т.к. плохо развитый эпидермис является воротами для проникновения инфекции и потенциальных токсинов из локально приложенных субстанций. Приобретение неполного барьера детьми преждевременно также м. вызывать дегидратацию, дисбаланс электролитов и плохую терморегуляцию. Хотя переход к земному существованию ex utero ускоряет программу эпидермальной дифференцировки, однако ребёнок, родившийся на 25-30 неделе нуждается в 2-4 неделях, чтобы сформировать функциональный барьер [1].
Существуют также множественные наследуемые нарушения epidermal permeability barrier (EPB), включая ichthyosis vulgaris, lamellar ichthyosis, X-linked ichthyosis и harlequin ichthyosis. Некоторые из этих заболеваний имеют под собой генетические дефекты, идентифицированные как мутации хорошо известных компонентов этого процесса [2]; однако, для большинства из них подлежащие генетические дефекты или плейотропны или картируются в областях, которые не содержат очевидных генов кандидатов.
Правда в последнее время достигнуты определенные успехи в понимании формирования EPB у мышей в качесиве модели. Мутантные мыши воспроизводят некоторые из этих фенотипических представлений, но они выявляют также легкие дефекты в вышестоящих частях путей, которые м. помочь в выявлении подлежащих генетических программ и механизмов.
Formation of the epidermal permeability
barrier
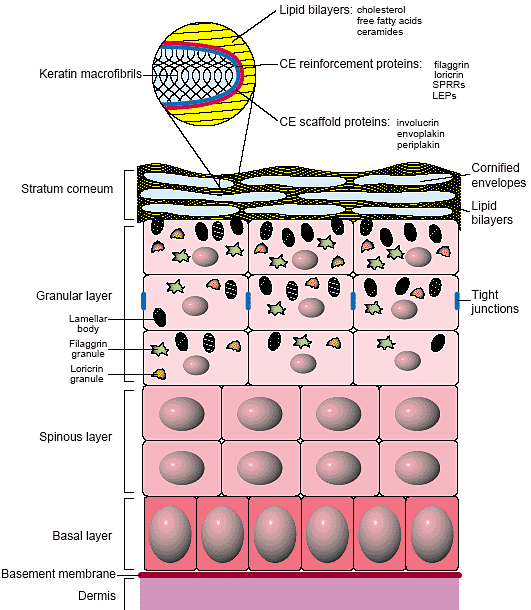
Эпидермис млекопитающих - стратифицированный эпителий - является само-обновляющейся тканью, состоящей из популяции митотически активных клеток в наиболее внутреннем базальном слое и их производных, которые перемещаются вверх к поверхности кожи согласно линейной программе окончательной дифференцировки. Этот процесс начинается, когда базальные клетки соответственно выходят из клеточного цикла, теряют слипчивость с базальной мембраной и начинают дифференцироваться. В промежуточных покрытых колючками (spinous) слоях клетки остаются транскрипционно активными, синтезирующими и собирающими прочную цитоскелетную сеть, которая создаёт механическую силу.
В верхнем гранулярном слое клетки уплощаются и деградируют своё внутриклеточное содержимое (включая ядра). Каждая клетка сохраняет кератиновые макрофибриллы и lipid-containing lamellar bodies (LBs), которые сливаются с плазматической мембраной. Постепенно собирается cornified envelope (CE) непосредственно под плазматической мембраной с помощью последовательного включения белков предшественников. Наконец, в наиболее наружном слое stratum corneum (SC), клетки становятся проницаемыми и приток кальция активирует transglutaminases для необратимого взаимного связывания белков CE, создавая
жёсткий, нерастворимый мешок, который окружает кератиновые волокна. CE служит в качестве поддержки (scaffold) для липидов, выдавливаемых из LBs, которые в свою очередь сливаются вместе с CEs, чтобы создавать барьер на поверхности кожи (Рис. 1) [3,4].
Имеющиеся данные подтверждают модель упорядоченной сборки CE с созданием инициальной белковой поддержки, с которой липиды поперечно связаны и с последующим усилением с помощью дополнительных структурных белков. Инициальная поддержка состоит из cytolinker plakin белков, envoplakin и periplakin, и структурного белка involucrin. В ответ на увеличение уровней Ca2+ активируется фермент transglutaminase 1 (Tg1), чтобы связать поперечно поддерживающие белки для образования непрерывного слоя.
Assembly of the epidermal permeability barrier. Schematic illustration of components (proteins and lipids) that contribute to the epidermal
barrier and where they are synthesized. Inset shows the assembly of the cornified envelope with the initial scaffold, lipid bilayers and the reinforcement proteins.
на внутренней поверхности плазматической мембраны. Следующей стадией сборки CE является слияние липиды (церамидов, свободных жирных кислот и холестерола), содержащих ламеллярных телец с плазматической мембраной, чтобы выдавить их содержимое во внеклеточное пространство.
Липидный бислой. Рис. с , где м. найти дополнительную информацию
Цепочки жирных кислот церамидов достаточно длинные, чтобы распространиться и в конечном итоге заместить плазматический липидный бислой. Церамиды ковалентно прикреплены к поддерживающим образованиям CE внутренне, и одновременно организуют внеклеточные липиды. Наконец, др. структурные белки, включая loricrin (который составляет ~80% CE), small proline-rich proteins (Sprr) и минорные количества др. белков, включая repetin и late envelope proteins (LEPs), поперечно связываются с поддерживающими образованиями, чтобы усилить инициальные структуры. Хотя большинство др. клеточных структур деградирует во время этого процесса, но keratin-filaggrin макрофибриллы оказываются поперечно связанными с CE посредством соединений с инициальной поддержкой и усиливающими белками [1] (Рис. 1).
Исторически, структуры. которые обеспечивают барьерную функцию, были описаны как 'bricks and mortar', с окончательно дифференцированными эпидермальнми клетками, формирующими кирпичики (bricks) и липиды, действующие как строительный раствор (mortar) [5]. Недавно продемонстрирована существенная роль межклеточных tight junctions (TJs) в эпидермальном барьере [6]. Др. межклеточные соединения, десмосомы, как полагают, играют роль в зарождении (nucleating) инициальных поддерживающих образовании CE и затем постепенно они исчезают с помощью упорядоченного процесса.
Assessing defects in epidermal permeability
barrier
Первоначальный прорыв произошел в области лучше всего видимых стадий индукции барьера во время эмбрионального развития у модельных животных. Группа Byrne's разработала качественный whole-mount dye permeability (WM-DP) подход для измерения первой стадии образования барьера. Этот подход показал, что EPB являются высоко упорядоченными во время развития с самостоятельными сайтами инициации, которые распространяются по эпидермису в виде движущегося фронта. Изменения в EPB , выявляемые с помощью WM-DP подхода коррелируют с ранее полученными данными, включая пониженную trans-epidermal water loss (TEWL),
презентацию зрелых CEs и слитые липиды LBs. Эти подходы для измерения накопления EPB в развитии позволили анализировать потери EPB у трансгенных мышей
[7].
Defects in catalyzing enzymes
Хотя мыши с дефицитом компонентов EPB обнаруживают дефекты в развитии или у новорожденных, эти животные показали удивительную способность к постнатальной компенсации. Напр., Tg1-дефицитные мыши погибают постнатально благодаря тяжёлой потере жидкости, обнаруживают проницаемость для краски и высокие уровни TEWL во время эмбрионального развития и у новорожденных. Если кожа неонатальных Tg1-/- трансплантируется мышам nude, то она накапливает утолщенные чешуйки, которые предупреждают потерю воды с поверхности кожи, снижая TEWL почти до нормального уровня [8].
Defects in lipid biosynthesis
В качестве иного примера, если доминантно-негативный retinoic acid receptor (RAR)a403 экспрессируется с промотора keratin 1 человека, то трансгенные мыши обнаруживают красную, лоснящуюся, липкую кожу при рождении с повышенной TEWL. Ультра структурный анализ выявил, что LBs образуются и сливаются с клеточной мембраной, но неспособны формировать мультиламеллярные структуры между клетками в SC. Хотя большинство keratin-1-RARa403 трансгенных животных становятся жертвами перинатальной летальности, некоторые затронутые мыши м. выживать, давая hyperkeratotic чешуйки [9].
Defects in protein precursors of the cornified
epithelium
В качестве третьего примера мыши, дефицитные по loricrin, главному структурному белку CE, обнаруживают задержку формирования EPB во время эмбрионального развития. Однако, loricrin-дефицитные новорожденные не обнаруживают каких-либо нарушений барьера. Молекулярный анализ показал, что происходит усиление активности др. белков, которые усиливают CE, включая Sprr
белки и repetin [10]. Сходные не летальные фенотипы выявлены у мышей, дефицитных по др. основным компонентам инициальных поддерживающих структур CE, включая envoplakin и involucrin [11,12].
Молекулярная природа компенсаторного механизма у loricrin-дефицитных мышей непосредственно сравнивалась с таковой у RARa403 трансгенных мышей (D Roop, personal communication). Такой подход необходимо расширить и на др. мутантных мышей с дефектами в EPB. Задача, чтобы понять, какие белки м. компенсировать отсутствие др. и является ли компенсация полной даже в условиях средовых стрессов. Первой ступенью д.б. анализ, какй из др. генов неправильно регулируется в отсутствие одного из основных структурных белков. Следующей ступенью д.б. тестирование функциональной компенсации с помощью анализа компаундных целенаправленных делеций. Основная проблема этой стратегии в том, что многие гены, кодирующие структурные белки CE, включая те, что кодируют loricrin, involucrin и repetin, и семейство Sprr и LEP генов, все картируются внутри области в 2.5 Mbp, известной как epidermal differentiation complex (EDC) [13,14]. Следовательно, их целенаправленного удаления невозможно достичь просто с помощью скрещиваний, чтобы получить компаундные гетерозиготы. Правда получение гнёздных делеций EDC м.б. внести определенную ясность.
Defects in intercellular junctions
Др. уровень сложности связан с функциональной ролью TJs в эпидермисе. Мыши дефицитные по claudin 1, трансмембранному компоненту TJs, обнаруживают классические симптомы дисфункции EPB, включая повышенную TEWL и потерю жидкостей тела. Однако, экспрессия маркёров дифференцировки и гистология эпидермиса и CEs выглядят нормально. В этих исследованиях применён новый анализ EPB путём инъекции трассирующей окраски низкого мол. веса подкожно, которая диффундирует через межклеточные пространства эпидермиса, до тех пор, пока не будет арестована в середине гранулярного слоя в TJs, которые становятся видимыми из-за окрашивания occludin. У claudin-1-дефицитных мышей TJs, по-видимому, формируются, но краска проходит через богатые occludin сайты [6]. TJs являются не просто барьером: они обнаруживают избирательность в отношении ионов и размеров и их барьерная функция варьирует существенно в зависимости от состава и соотношения в смеси молекул claudin в индивидуальных спаренных полосках TJ [15]. Если claudin 6 эктопически экспрессируется с промотора involucrin в верхних spinous и гранулярном слоях, то мыши имеют дефекты EPB, как показывает WM-DP подход и измерения TEWL. Turksen and Troy [16] изучали экспрессию белков, участвующих в образовании CE, и нашли некоторые изменения в уровнях процессинга filaggrin и разных членов семейства Sprr, указывающих на то, что имеется некий механизм обратной связи между этими двумя компонентами EPB. Интересно бы определить, существует ли какой-либо компенсаторный механизм когда трансгенная involucrin-claudin-6 и claudin-1-дефицитная кожа трансплантируется мышам.
Десмосомы, как полагают, зачинают (nucleate)инициальную сборку поддерживающих образований CE and и идентифицируются как поперечно связывающие компоненты CE. В SC десмосомы прогрессивно деградируют с помощью не охарактеризованного ещё механизма. Когда соотношение кадхеринов десмосом изменено с помощью экспрессирующегося эктопически desmoglein 3 в spinous слоях эпидермиса, то десмосомы преждевременно растворяются и мыши теряют функцию барьера как новорожденные [17]. Мыши с целенаправленным устранением гена десмосомального кадхерина desmocollin 1 устанавливают барьер in utero и имеют нормального вида CE при рождении. Однако, вообще-то из-за вторичного эффекта эпидермальной ломкости, новорожденные обнаруживают многочисленные области локальной потери барьерной функции [18]. Необходимы дальнейшие исследования роли специфических белков десмосом в инициации сборки CE и функции исчезновения десмосом в установлении барьера.
Defects in transcriptional regulators
Регуляторы, которые, как полагают, расположены выше пути контроля образования EPB также обнаруживают некоторую степень компенсации, д.б. пригодны для анализа того, как EPB формируются и как это м. противодействовать потере. Напр., антенатальная материнская терапия глюкокортикоидами снижает смертность у детей перед рождением за счёт ускорения продукции surfactants в лёгких и созревания EPB [19]. Сходным образом фармакологические дозы глюкокортикоидов вызывают ускоренное созревание эпидермиса у мышей и крыс [7,20]. Напротив, дефицитные по глюкокортикоиду эмбрионы имеют задержанные в развитии EPB что демонстрируется по созреванию SC и LB. При рождении, однако, дефицитные по глюкокортикоиду эмбрионы обладают нормальными EPB [21]. Молекулярная природа глюкокортикоидных мишеней и что м. функционально компенсировать их потерю во время развития остаются неизвестными.
В качестве др. примера м. служить транскрипционный фактор kruppel-like factor 4 (KLF4), экспрессирующийся в дифференцирующихся слоях эпидермиса. Мыши KLF4-/- погибают вскоре после рождения из-за потери функции EPB, как показывают измерения с помощью WM-DP, и быстрой потери жидкости тела. CEs являются незрелыми, а секретируемые липиды неспособны формировать бислои, собственно соединяющие CEs. Дефект EPB не м.б. исправлен с помощью трансплантации KLF4-/- неонатальной кожи [22]. Напротив экспрессия KLF4 с промотора keratin5 (K5-KLF4) на более ранних стадиях развития обычно ускоряет приобретение барьера, что проявляется в непроницаемости для краски, в экспрессии маркёров терминальной дифференцировки и в сборке зрелых CEs [23].
Гены, кодирующие компоненты CE были идентифицированы благодаря неправильной регуляции KLF4-дефицитной кожи [22]. Ведутся исследования по выявлению генов. которые обратным образом регулируются у KLF4-/- и K5-KLF4 трансгенных мышей, которые м. представлять собой непосредственные мишени для KLF4 в приобретении EPB. Первые исследования показали, что kruppel-подобные транскрипционные факторы и глюкокортикоидные рецепторы м. действовать синергично, способствуя суперактивации транскрипции [24]. Сравнивая результаты молекулярных профилей мышей с модулированными уровнями KLF4 и glucocorticoids в дальнейшем м. будет выяснить общий путь у этих двух факторов
.
Conclusions and future prospects
Имеющиеся данные подтверждают модель ступенчато организованной сборки CE в нормальных условиях. Однако, др. теории, включая 'dustbin hypothesis' или модифицированную 'precursor availability hypothesis' м. описать, что происходит при генетических или средовых стрессовых ситуациях. Эти модели отличаются от модели организованной сборки в предположении, что transglutaminase ферменты д. использоваться независимо от того какие предшественники доступны для формирования CE [25,26]. Гипотеза мусорного ящика (dustbin) или гипотеза доступности предшественников м. объяснить, почему отсутствие определенных предшественников CE м.б. частично компенсировано за счёт усиления активности др. компонентов CE и почему определенные белки, такие как Sprr и LEP, неправильно регулируются, когда эпидермис реагирует на потерю барьера.
Сходного типа анализ подлежащих генетических дефектов у людей с наследственными нарушениями cornification представлен в обзоре Steinert и др. [27]. Предстоит исследование огромного количества мутаций у мышей с дефектами эпидермальной проницаемости у новорожденных, взрослых и при трансплантациях кусочков взрослой кожи (Table 1). Степень избыточности и сложности в становлении EPB замечательна, если учесть что эти механизмы лишь недавно сформировались, когда организмы перешли к наземному существованию.