Установлено, что в большинcтве случаев первые деления дробления генерируют один бластомер, чье потомство будет восполнять эмбриональную часть бластоциста (полярную трофэктодерму и более глубокие клетки внутренней клеточной массы) и др.. чьё потомство будет давать неэмбриональную часть (mural трофэтодерма и наиболее поверхностная часть внутренней клеточной массы)
. Дальнейшие эксперименты по отслеживанию клонов прояснили эти взаимоотношения и показали, что у 4-клеточных нормально развивающихся эмбрионов бластомеры не только обладают специфической судьбой, но и м. также отличаться своим онтогенетическим потенциалом
. Установлено, что в нормальных условиях первое деление дробления обычно проходит в близи этого полюса
. Второй маркер первого деления дробления, место вхождения спермия
, отражает расположение веретена относительно несколько уплощенной зиготны при оплодотворении
. Возникает вопрос, возникает ли пространственная организация зиготы по отношению к анимальному полюсу или из-за того, что ориентация дробления м. отражать случайное место проникновения спермия, и само дробление, которое влияет на последующее развитие на эмбриональную-не-эмбриональную ось.
Разработан улучшенный метод для аккуратного трехмерного изучения ориентации первого деления дробления в случайной популяции зигот. Оценивалось это с помощью time-lapse наблюдений первого деления дробления. Кроме того положение плоскости первого деления определяли в отношении пронуклеусов (и polar body (PB)) во вновь полученной трансгенной линии (CAG:H2B-EGFP), в которой хроматин был мечен с помощью экспрессии green fluorescent protein (GFP)-меченного гистона histone H2B (ref. 10; Fig. 1a; Supplementary Movie 1). Было устанолвено, что женский пронуклеус находился ближе к PB, чем мужской пронуклеус в 87.5% (n = 32) зигот (categories I-III; Fig. 1b) и что у большинства зигот (category I, 62.5%) оба пронуклеуса были выровнены по PB, а борозда дробления формировалась внутри 30
о этой плоскости. У 12.5% зигот (category II), пронуклеусы помещались по сравнению с PB между 30
о и 90
о, а плоскость дробления смещалась в той же самой степени. В двух оставшихся категориях дробление происходило внутри 30-90
о с PBs выровненными по пронуклеусам (category III) и внутри 30
о с PBs не выровненными с пронуклеусами (category IV). Мы наблюдали, что это наиболее частое расположение дифференциально конденсированных
11 женских и мужских пронуклеусов с PB сохраняется и в прометафазе. Иммуноокрашивание фиксированных зигот, чтобы выявить CpG-метилированнй женских хроматин, показало, что по сравнению с предыдущими исследованиями
11,12 и в противоположность недавнему предположению
13, родительские наборы хромосом не смешиваются и принимают расположение, согласное с нашими time-lapse исследованиями, что они сохраняются и в дочерних бластомерах (Fig. 1c). Чтобы охарактеризовать ориентацию плоскости деления далее мы осуществили time-lapse differential interference contrast (DIC) микроскопию зигот разных линий мышей дикого типа (Fig. 1d). Наблюдения со множественных точек гарантировало, что мы смогли проследить деление по отношению к маркерам в трех измерениях в 96% всех случайно расположенных зигот. Мы нашли, что начало формирования борозды дробления внутри 30
о плоскости PB у 70% зигот в 14 раз выше, чем в латеральном положении между
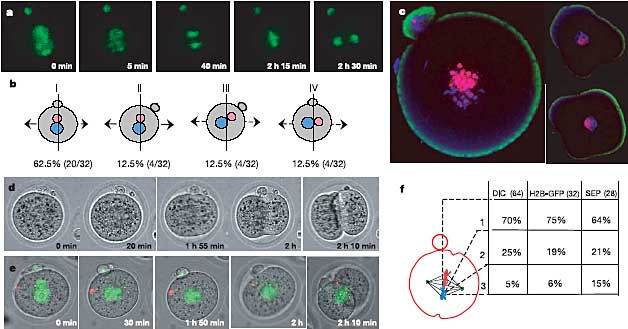 Figure 1 Orientation of the first cleavage division. a, One zygote expressing H2B-EGFP from 32 embryos observed by time-lapse imaging, to note the positions of maternal and paternal chromosomes before chromosome condensation, during prometaphase, anaphase and cytokinesis. b, Four cleavage categories of zygotes from a (vertical line, cleavage plane; horizontal arrows, direction of cell separation): I, male and female pronuclei aligned within 308 of PB; cleavage passed through this plane; II, male and female pronuclei displaced from PB by 30-908; cleavage through plane of pronuclei; III, pronuclei aligned within 308 of PB; cleavage displaced from this plane by 30-908; IV, pronuclei not aligned with PB; cleavage within 308 of the PB. c, Zygotes fixed and stained to reveal maternal (CpG methylated chromatin, pink) and paternal chromatin (blue). Left, prometaphase zygote; right, optical sections through separate two-cell blastomeres. Parental chromosomes do not mix during the first cleavage and still occupy distinct territories at the two-cell stage. d, DIC time-lapse series of zygote with cleavage at the site of the PB. e, Time-lapse series of H2B-EGFP (green) labelled zygote with red fluorescent bead at sperm entry site. a, c-e, The diameter of the zygote is approximately 80mm. f, Proportions of zygotes with cleavage plane within 308 (1), 30-608 (2) and 60-908 (3) of the PB in a DIC time-lapse series of 64, a series of 32 to follow H2B-EGFP, and a series of 28 to assess cleavage in relation to the fluorescent bead marking sperm entry.
Figure 1 Orientation of the first cleavage division. a, One zygote expressing H2B-EGFP from 32 embryos observed by time-lapse imaging, to note the positions of maternal and paternal chromosomes before chromosome condensation, during prometaphase, anaphase and cytokinesis. b, Four cleavage categories of zygotes from a (vertical line, cleavage plane; horizontal arrows, direction of cell separation): I, male and female pronuclei aligned within 308 of PB; cleavage passed through this plane; II, male and female pronuclei displaced from PB by 30-908; cleavage through plane of pronuclei; III, pronuclei aligned within 308 of PB; cleavage displaced from this plane by 30-908; IV, pronuclei not aligned with PB; cleavage within 308 of the PB. c, Zygotes fixed and stained to reveal maternal (CpG methylated chromatin, pink) and paternal chromatin (blue). Left, prometaphase zygote; right, optical sections through separate two-cell blastomeres. Parental chromosomes do not mix during the first cleavage and still occupy distinct territories at the two-cell stage. d, DIC time-lapse series of zygote with cleavage at the site of the PB. e, Time-lapse series of H2B-EGFP (green) labelled zygote with red fluorescent bead at sperm entry site. a, c-e, The diameter of the zygote is approximately 80mm. f, Proportions of zygotes with cleavage plane within 308 (1), 30-608 (2) and 60-908 (3) of the PB in a DIC time-lapse series of 64, a series of 32 to follow H2B-EGFP, and a series of 28 to assess cleavage in relation to the fluorescent bead marking sperm entry.
60
о и 90
о PB (Fig. 1a, f). У тех зигот, у который она локализовалась в медиальном положении (25%), PB обычно оказывался втянут в борозду и в последующем входит в состав более чем 90% эмбрионов. Т.о., первое деление дробления имеет тенденцию проходить вблизи области PB, в плоскости, вдоль которой располагаются родительские геномы. Мы также проследили флюоресценцию как GFP-меченного H2B, так и глобулы, отмечающей место проникновения спермия с помощью time-lapse наблюдений (Fig. 1e и Supplementary Movie 2). Это показало, что дробление проходит внутри 30
о от места проникновения спермия у подавляющего числа зигот (Fig. 1e, f). Кроме того мы предприняли небольшую серию трансплантационных экспериментов, чтобы изменить место анимального полюса и чтобы понаблюдать с помощью time-lapse imaging, будет ли дробление обнаруживать тенденцию к прохождению черех это новое место. Почти у всех (6 из 7) эмбрионов первое деление дробления происходило внутри 30
о вновь позиционированного PB (Supplementary Movie 3). Эти исследования на живых эмбрионах подтвердили более ранние работы, показавшие, что имеется склонность прохождения первого деления дробления вблизи прикрепленного PB
7-9 и места проникновения спермия
1,9. Недавние исследования показали, что дробление из двух-клеточного эмбриона бластомера, который наследует PB, также имеет тенденцию проходить вблизи PB (ref. 5).
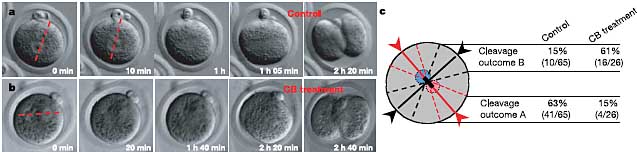 Figure 2 Cytochalasin treatment before pronuclei migration changes the orientation of the first cleavage division. a, b, Time-lapse series of control (a) and cytochalasin-treated (b)
zygotes. The diameter of the zygotes is approximately 80mm. c, Diagram to show the plane represented by extrapolation of final trajectories of two pronuclei (solid red line).Cleavage within 308 of this line is represented as 'outcome A'. The solid black line represents cleavage 908 to this final path of pronuclear migration. Cleavage within 308 of
this is referred to as 'outcome B'. The relative proportions of cleavages that followed these two outcomes in control and cytochalasin-treated embryos are shown.
Figure 2 Cytochalasin treatment before pronuclei migration changes the orientation of the first cleavage division. a, b, Time-lapse series of control (a) and cytochalasin-treated (b)
zygotes. The diameter of the zygotes is approximately 80mm. c, Diagram to show the plane represented by extrapolation of final trajectories of two pronuclei (solid red line).Cleavage within 308 of this line is represented as 'outcome A'. The solid black line represents cleavage 908 to this final path of pronuclear migration. Cleavage within 308 of
this is referred to as 'outcome B'. The relative proportions of cleavages that followed these two outcomes in control and cytochalasin-treated embryos are shown.
В приведенных выше экспериментах мы не отбирали и не исключали эмбрионы, а проанализировали 96% от всех учтенных зигот с множественных фокальных плоскостей в каждый момент времени. Напротив, в недавнем исследовании, которое изучало деление дробления по отношению к PB в одной фокальной плоскости, было выявлено более 50% зигот, у которых дробление оказалось чувствительным к позиции наблюдения
13. Более того, чтобы проверить дробление в отношении пронуклеусов. те же самые авт выбирали эмбрионы, у которых пронуклеусы располагались в одной и той же фокальной плоскости
13. Это указывает на то, что их данные м. обнаруживать склонность к отбору специфических групп эмбрионов для анализа. Это м. б. объяснить, почему их заключение, что первое деление дробления не связано с PB, а более того совпадает с плоскостью между apposing пронуклеусами, отличается от четырех-мерных исследований, описанных в данной работе. Такая склонность м. возникать из-за положения зигот
 Figure 3 Relationship between final position of pronuclei, long axis of zygote and first cleavage in experimentally elongated zygotes. a, b, PB at the end of the long axis; c, d, PB
at the end of the short axis. a, c, Proportions of zygotes in which alignment of pronuclei was within 308 (left), 61-908 (middle) and 31-608 (right) of the long axis in a time-lapse series of 51 zygotes. b, d, Proportions of zygotes that showed a cleavage plane within 308
(top), 31-608 (middle) and 61-908 (bottom) of the short axis. e, Example of a field of experimentally elongated embryos at the beginning (20 h after hCG injection, left) and end (34 h 10 min after hCG injection, right) of a time-lapse series. f, One example of a zygote at five time points. The ratio between the short and long axis of this zygote was 0.83. Time points: at 0 min (beginning of the experiment 18 h after hCG injection) both pronuclei are still apart from each other; at 2 h 15 min, both pronuclei have met in the centre of the zygote; at 13 h 55 min, the last frame in which pronuclei can still be seen; at 15 h 45 min, elongated zygotes are preparing for cleavage; at 15 h 50 min,cytokinesis occurs. The diameter of the not elongated embryos is approximately 80mm.
Figure 3 Relationship between final position of pronuclei, long axis of zygote and first cleavage in experimentally elongated zygotes. a, b, PB at the end of the long axis; c, d, PB
at the end of the short axis. a, c, Proportions of zygotes in which alignment of pronuclei was within 308 (left), 61-908 (middle) and 31-608 (right) of the long axis in a time-lapse series of 51 zygotes. b, d, Proportions of zygotes that showed a cleavage plane within 308
(top), 31-608 (middle) and 61-908 (bottom) of the short axis. e, Example of a field of experimentally elongated embryos at the beginning (20 h after hCG injection, left) and end (34 h 10 min after hCG injection, right) of a time-lapse series. f, One example of a zygote at five time points. The ratio between the short and long axis of this zygote was 0.83. Time points: at 0 min (beginning of the experiment 18 h after hCG injection) both pronuclei are still apart from each other; at 2 h 15 min, both pronuclei have met in the centre of the zygote; at 13 h 55 min, the last frame in which pronuclei can still be seen; at 15 h 45 min, elongated zygotes are preparing for cleavage; at 15 h 50 min,cytokinesis occurs. The diameter of the not elongated embryos is approximately 80mm.
в культуральных чашках из-за их формы. Это особенно важно, т.к. форма зигот м. отменять любое влияние области, маркируемой с помощью PB в отношении ориентации первого деления дробления
9. Если бы мы отбирали зиготы определенной формы, то смогли бы мы также наблюдать разные взаимоотношения первого деления дробления к позиции пронуклеусов?
Обработка зигот cytochalasin в концентрациях, которые разрушают актиновый цитоскелет, д. мешать эффекту естественного изменения формы, индуцируемого оплодотворением, чтобы повлиять на положение борозды дробления
9. Мы поэтому изучали дробление у эмбрионов, обработанных cytochalasin в течение 4 ч в концентрации, которая деполимеризует актиновый цитоскелет примерно за 6 ч до дробления. В противоположность контрольным зиготам (Fig. 2, outcome A; Supplementary Movie 4), плоскость дробления у обработанных cytochalasin зигот (Fig. 2, outcome B; Supplementary Movie 5) находится внутри 30
о плоскости, ортогональной по отношению к финальной траектории пронуклеусов. Т.о., только у обработанных эмбрионов мы наблюдали конечный результат, сходный с тем, что был описан ранее
13. В самом деле, мы отметили, что эти эксперименты, которые привели к заключению, что плоскость дробления находится между располагющимися рядом трансплантированными пронуклеусами
13, только если зиготы были обработаны cytochalasin, для осуществления необходимых манипуляций. Такой обработки достаточно, чтобы повлиять на ориентацию дробления. Мы установили, что время обработки cytochalasin является критическим для конечного результата. Если он используется рано в ходе миграции пронуклеусов, т.к. это необходимо для трансплантации пронуклеусов, то дробление более не зависит от положения PB (44% делений внутри 30
о PB, 36% между 30
о и 60
о и 20% между 60
о и 90
о; n = 25). Если зиготы были обработаны cytochalasin, то мигрировали внутрь, как в наших экспериментах по трансплантации цитоплазмы из области PB, а большинство дроблений (73%, 16 из 22) уважало PB.
Наши предварительные исследования показали, что если зигота экспериментально 'flattened', то дробление проходит по оси нового типа, которая отменяет эффекты анимального полюса и места проникновения спермия
9. М. ли такое изменение формы повлиять на расположение пронуклеусов в отношении длинной и короткой осей зиготы и на их взаимоотношения с плоскостью дробления? Мы меняли форму зигот, помещая их в микропипетку и высвобождая в sodium alginate, так что PB располагался на конце или длинной или короткой оси. Мы установили, что независимо от позиции PB, пронуклеусы стремятся расположиться вдоль новой длинной оси зиготы (Fig. 3). У большинства зигот (59%, 30 из 51)
пронуклеусы располагались внутри 30
о длинной оси и только 16% (8 из 51) имели пронуклеусы внутри 30
о короткой оси. Дробление происходит внутри 30
о короткой оси у 92% (47 из 51) от всех зигот. Т.о., плоскость деления проходит в плоскости между двумя apposing пронуклеусами у зигот, чья форма позволят избегнуть влияния со стороны PB маркированной области на положение плоскости дробления. Это контрастирует с огромным большинством не подвергнутых воздействию зигот, у которых пронуклеусы располагаются вдоль плоскости первого деления дробления. Положение пронуклеусов, по-видимому, не зависит от плоскости дробления (Fig. 3). Всё это указывает на то, что клеточная форма скорее. чем положение пронуклеусов влияет на плоскость дробления у удлиненных эмбрионов. Т.о., мы не м. подтвердить заключение, сделанное ранее
13, что зиготы всегда делятся в соответствии плоскости, разделяющей два apposing пронуклеуса. Мы также выявили подобные паттерны дробления только внутри отобранной группы эмбрионов, которые или обработаны cytochalasin до миграции пронуклеусов, или испытывали изменения формы, так что пронуклеусы оказывались лежащими вдоль длинной оси.
Эта способность манипулировать зиготами, чтобы делить воспроизводимо таким альтернативным образом. позволила нам определить последовательность развития эмбриональной-не-эмбриональной оси бластоциста. Мы изменяли форму зиготы и культивировали эмбрионов в среде. содержащей sodium alginate вплоть до двухклеточной стадии. Эмбрионы, которые делились с PB на одном конце длинной оси, затем удаляли из solidified среды и их бластомеры метили разными липофильными красками (DiI или DiD, Molecular Probes). Мы позволяли таким меченным эмбрионам развиваться в бластоцисты и определяли положение клонов в отношении полости бластоциста с помощью конфокального sectioning
(Fig. 4). У зигот, чья форма не была изменена, два соотв. клона обнаруживали тенденцию занимать эмбриональную (полярная трофэктодерма и и более глубокие клетки внутренней клеточной массы) и не-эмброиональную (mural trophectoderm) части бластоциста.
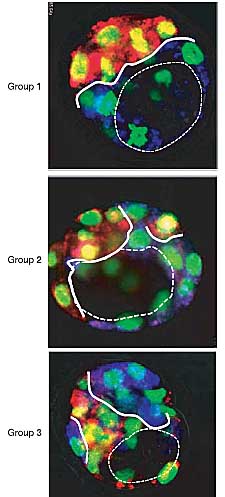 Figure 4 Outcome of changing orientation of first cleavage on the orientation of blastocyst embryonic-abembryonic axis. Zygotes were elongated such that PB was at one end of the
long axis. On their division to the two-cell stage, one blastomere was labelled with DiI (red) and other with DiD (blue). At the blastocyst stage, a total of 20 embryos were obtained and classified according to the extent to which cells derived from one blastomere comprising
mainly the embryonic part crossed the embryonic-abembryonic boundary zone (a region approximately one cell deep and parallel to the roof of the blastocyst cavity) into the abembryonic part and vice versa. In group 1, up to two cells crossed the boundary zone; in group 2, three cells crossed the boundary zone; and in group 3, more than three cells
crossed the boundary zone. Micrographs represent individual optical sections midway through the embryo that show the cavity (thin dashed line) and clonal boundary zone (thick solid line). The diameter of the blastocysts is approximately 80mm.
Figure 4 Outcome of changing orientation of first cleavage on the orientation of blastocyst embryonic-abembryonic axis. Zygotes were elongated such that PB was at one end of the
long axis. On their division to the two-cell stage, one blastomere was labelled with DiI (red) and other with DiD (blue). At the blastocyst stage, a total of 20 embryos were obtained and classified according to the extent to which cells derived from one blastomere comprising
mainly the embryonic part crossed the embryonic-abembryonic boundary zone (a region approximately one cell deep and parallel to the roof of the blastocyst cavity) into the abembryonic part and vice versa. In group 1, up to two cells crossed the boundary zone; in group 2, three cells crossed the boundary zone; and in group 3, more than three cells
crossed the boundary zone. Micrographs represent individual optical sections midway through the embryo that show the cavity (thin dashed line) and clonal boundary zone (thick solid line). The diameter of the blastocysts is approximately 80mm.
Количество клеток от каждого клона в пограничной зоне, lying one cell
deep и параллельно поверхности бластоцеля
3, м. варьировать. Чтобы сравнить распределение потомства от обойих бластомеров двухклеточного эмброиона. мы оценивали степень, с которой клоны распространялись за пределы пограничной зоны у 20 меченных бластоцист, измененных так, что PB оказывался на одном из концов длинной оси после первого дробления (Fig. 4 and Supplementary Fig. 1). Или не было или очень мало клеток (до двух, примерно 3-9% от общего среднего количества клеток (n = 22.25) такие бластоцистя относили к группе 1; если три клетки пересекали границу, то бластоцисты относили к группе 2; и если более трех, то бластоцисты относили к группе 3. В группе 1, у 85% бластоцистов (17 из 20) две или менее клеток от преимущественно эмбрионального клона оказывалось за пределами пограничной зоны в не-эмбриональной части (Fig. 4) и у 65% (13 из 20) из тех же самых бластоцистов две или меньше клеток преимущественно не-эмбрионального клона оказывалось в эмбриональной части (Fig. 4). В группе 2, у 15% (3 из 20) бластоцистов три клетки из преимущественно эмбрионального клона локализовались в не-эмбриональной части; в др. направлении пропорция составляла 20% (4 of 20 бластоцистов). В группе 3 не было (0 из 20) бластоцистов, у которых три или более клеток из преимущественно эмбрионального клона присутствовало в не-эмбриональной части. Только 15% (3 of 20) из бластоцистов этой группц обнаруживало преимущественно не-эмбриональный клон, распространяющийся в эмбриональную часть. Т.о., у большинства эмбрионов, большая часть потомства от бластомеров двух-клеточного эмбриона остается или в эмбриональной или не-эмбюриональной части бластоциста. Сходная пропорция эмбрионов обнаруживала такие взаимоотношения и в предыдущих исследованиях non-manipulated эмбрионов, у которых дробления имеют тенденцию проходить вблизи анимального полюса
3. Мы также наблюдали относительно небольшое перемешивание клеток двух клонов от бластоцистов ранней стадии
14 (Supplementary Fig. 1).
Наше исследование дополняет ранее полученные доказательства взаимоотношений между первым делением дробления и областью соседствующей с PB5-9. Однако, это взаимоотношение области анимального полюса с ориентацией дробления м.б. преодолено с помощью эффектов формы на зиготу. как это было продемонстрировано ранее [9]. Мы установили, что независимо от ориентации деления дробления в отношении пронуклеусов или PB, у большинства эмбрионов один из двух-клеточных бластомеров занимает эмбриональную часть бластоциста, а др. занимает вне-эмбриональную часть.
Т.о., образование оси эмбрион-внеэмбриональная часть имеет тенденцию возникать с учетом плоскости первого деления дробления не зависимо от того, проходит она или нет вблизи анимального полюса. Это указывает ли это на то, что организация яйцеклетоки относительно анимального полюса не влияет на развитие всего остального кроме обычно скоординированного первого деления дробления? Недавние эксперименты подтвердили, что паттерны ранних делений дробления разделяют бластомеры не только с разными судьбами. но и с отличающимися онтогенетическими свойствами5,6. Это связано как с их специфической позицией в эмбрионе относительно анимального полюса, так и различий по времени делений этих клеток. Т.о., существование пространственно распределенных компонентов яйца работает сочетано с факторами, регулирующими клеточные деления, чтобы влияние на развитие не было обесценено.
Сайт создан в системе
uCoz