Myosin 3A transgene expression produces abnormal actin filament bundles in transgenic Xenopus laevis rod photoreceptors
J.Lin-Jones, E.Parker, M.Wu, A.Dose, B.Burnside J.Cell Sci. - 2004.- Vol. 117, No 24, P. 58925-5834
Экспрессировали Myo3A, меченный GFP, в палочковидных фоторецепторах трансгенных Xenopus, используя модифицированный промотор опсина. Меченнй интактный Myo3A локализуется в calycal отростках палочек. Трансгенные фоторецепторы палочки давали аномально большие calycal отростки и затем дегенерировали. Экспрессия модифицированного Myo3A показала, что локализация calycal отростков нуждается в активном моторном домене и хвостовом домене. В отдельности экспрессируемый хвостовой домен локализуется в актиновых пучках вдоль всей длины внутреннего сегмента скорее, чем на дистальном конце. Эта локализация хвостового домена нуждается в консервативном С-терминальном домене, обладающим актин-связывающим мотивом. Полученные данные указывают на то, что Myo3A играет роль в морфогенезе и поддержании calycal отростков фоторецепторов позвоночных.
Reyna I. Martinez-De Luna and Heithem M. El-Hodiri (elh@ccri.net ) The Xenopus ortholog of the nuclear hormone receptor Nr2e3 is primarily expressed in developing photoreceptors Int. J. Dev. Biol. 51: 235 - 240 (2007)
doi: 10.1387/ijdb.062236rm
Nr2e3 является рецептором ядерного гормона, который участвует в дифференцировке палочковидных фоторецепторов. Ген Nr2e3 ранее был идентифицирован у человека, мыши, рыбок данио и эмбрионов кур. У всех видов экспрессия Nr2e3 ограничена сетчаткой и как полагают, выполняет роль в спецификации и поддержании фоторецепторов палочек. Описывается идентификация и характеристика Xenopus Nr2e3. Установлено, что Nr2e3 преимущественно экспрессируется в развивающихся фоторецепторах палочках. В отличие от др. видов, Nr2e3 у лягушек экспрессируется также в хорде и шишковидной железе во время развития Xenopus laevis.
Неконвенциональные миозины класса III являются зависимыми от актиновых филамент моторными белками с N-терминальным киназным доменом (Kalhammer, Bahler, 2000). Эти миозины играют роль в в функции сенсорных клеток. Первый член семейства миозинов III ninaC клонирован у Drosophila/ он экспрессируется исключительно в зрительных фоторецепторах. Мутации в ninaC вызывают дефекты фототрансдукции, а также дегенерацию фоторецепторов, зависимую от света и возраста. у позвоночных, два гена миозина III, Myo3A и Myo3B экспрессируются в фоторецепторах сетчатки, а Myo2A экспрессируется также в ухе (Dose et al., 2003; Walsh et al., 2002). Недавно установлено, что мутации MYO3A человека ответственны за прогрессирующую несиндромальную глухоту (Walsh et al., 2002).
В фоторецепторах мух и позвоночных Myo3 ассоциирует с отростками, похожими на микроворсинки, содержащими в сердцевине актиновые филаменты. Ген ninaC Drosophila кодирует две альтернативно спласируемые изоформы (р132 и р174). Хотя обе экспрессируются в фоторецепторах , но р132 обнаруживается в клеточном теле, тогда как р174 в рабдомерах, в фотопигмент несущих микроворсинках, которые являются функциональным эквивалентом наружных сегментов фоторецепторов позвоночных. У ninaC мутантов организация стержневых актиновых филамент рабдомерных микроворсинок нарушена. В сетчатке сев. американских полосатых рыбок (striped bass) Myo3A локализуется в дистальных концах пучков актиновых филамент (Dose et al., 2003), которые формируют обойму (cage) под клеточной мембраной и распространяются от наружной ограничивающей мембраны через внутренний сегмент к кончикам подобных микроворсинкам calycal отростков, которые охватывают в виде бокала наружный сегмент (Nagle et al., 1986). Считается, что calycal отростки являются почти повсеместно распространенными в фоторецепторах позвоночных, их функция неизвестна.
Собранные в пучки актиновые филаменты внутреннего сегмента фоторецепторов позвоночных ориентированы своими минус-концами в направлении ядра и плюс концами в направлении дистальных кончиков calycal отростков (O'Connor, Burnside, 1981). Т.к. Myo3a является направленным к плюс-концам мотором (Komaba et al., 2003), то м. ожидать, что он транслоцируется дистально вдоль пучков актиновых филамент внутреннего сегмента в направлении плюс концов и , следовательно, накапливается в на дистальных концах филоподий (Erickson et al., 2003), указывая тем самым что в этой гетерологической системе Myo3A д. транслоцироваться к плюс концам актиновых филамент, образующих сердцевину филоподий.
Хвостовые области разных классов миозинов сильно отличаются и в целом, как полагают, специализированы к обеспечению тип-специфической субклеточной локализации и/или предназначены для самостоятельных функций разных миозинов. Myo3A имеет два законсервированных домена в своей хвостовой области: 3 tail homology damains I and II (3THDI и II) (dose et al., 2003). 3THDI состоит из ~58 аминокислотных последовательностей у всех миозинов класса III позвоночных, тогда как 3THDII из 22 аминокислот обнаруживается только на С-конце миозинов IIIAs позвоночных. Ранее было установлено, что дистальное накопление временно экспрессируемого Myo3A bass в кончиках филоподий клеток HeLa нуждается в присутствии как активного мотора, так и терминального 3THDII домена (Erickson et al., 2003). Этот 3THDII мотив обладает актин-связывающим доменом, сходным с тем, что обнаружен в гладкомышечной миозиновой легкой цепи киназы (Erickson et al., 2003).
В данном исследовании идентифицированы элементы Myo3A, необходимые для локализации в дистальной части внутреннего сегмента фоторецепторов и calycal отростках, используя трансгенных головастиков Xenopus laevis, у которых экспрессия ограничена палочковидными фоторецепторами с помощью промотора палочкового опсина Xenopus. Установлено,
что экспрессия трансгена Myo3A в палочках ведет к образованию аномально больших calycal отростков и к последующей дегенерации палочек. Это указывает на то, что Myo3A играет роль в морфогенезе и/или поддержании calycal отростков палочковидных фоторецепторов и актинового цитоскелета внутреннего сегмента фоторецепторов.
 E.Parker, M.Wu, A.Dose, B.Burnside
E.Parker, M.Wu, A.Dose, B.Burnside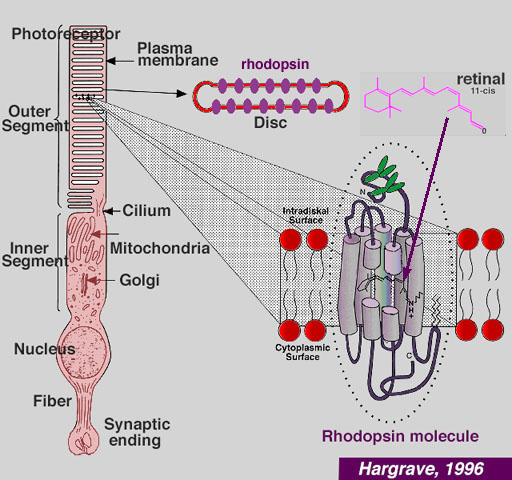
){kind=link}