Cleavage furrow positioning  The Journal of Cell Biology | Volume 164, Number 3, 2004 | |
|
To complete the cell cycle, the cleavage furrow draws the
plasma membrane toward the cell center, pinching the
cytoplasm into two lobes that are subsequently separated
into two cells. The position of the cleavage furrow is induced
by the mitotic spindle during early anaphase. Although
the mechanism of cleavage furrow positioning is not
understood at a molecular level, recent results suggest
that it might be mediated by local relief from the inhibitory
effects of microtubules.
|
В клетках животных механические силы для клеточного деления генерируются с помощью myosin II ,т.к. он транслоцирует актиновые филаменты внутри контрактильного кольца. Контрактильное кольцо собирается в клеточном кортексе в богатом актином слое, расположенном рядом с клеточной мембраной, в положении, которое предопределяется с помощью митотического веретена во время анафазы. Supramolecular control of furrow formation Хромосомы, как было установлено, несущественны для цитокинеза (Rappaport, 1996; Zhang and Nicklas, 1996; Bucciarelli et al., 2003; Dekens et al., 2003). Центросомы м.б. устранены или генетически разрушены без нарушения цитокинеза (Khodjakov and Rieder, 2001; Megraw et al., 2001). Хотя, как полагают, хромосомы и центросомы несущественны, они м. влиять на процесс, если присутствуют (Piel et al., 2001). Микротрубочки веретена деполимеризуются во время метафазы или очень ранней анафазы, предупреждая образования борозды дробления, указывая тем самым, что микротрубочки существенны (Hamaguchi, 1975). Однако, образование борозды м. происходить, если митотическое веретено деполимеризуется позднее в анафазе, но до начала проникновения (ingression) (Hamaguchi, 1975). Т.о., микротрубочки митотического веретена необходимы для индукции образования борозды, но они не per se необходимы для ingression.
Физические манипуляции с эмбриональными клетками включают изменение положения веретена по отношению к клеточному кортексу, деформации клеточной формы и удаление частей веретена. Напр., классический "torus experiment", в котором два веретена в общей цитоплазме индуцируют дополнительную борозду, указывает на то, что оппозитные звёздочки достаточны для индукции борозды (Rappaport, 1961). Кроме того, изменение позиции веретена во время анафазы приводит к возникновению множественных борозд, чьё положение диктуется веретеном (Rappaport, 1985). Эти исследования привели к созданию astral stimulation модели (Fig. 1 A; Rappaport, 1996). Согласно этой концепции астральные микротрубочки обеспечивают стимулы для деления, которым м.б., напр., фактор, который транспортируется вдоль астральных микротрубочек. Эта модель предполагает, что т.к. экваториальный кортекс испытывает влияние со стороны астральных микротрубочек от двух полюсов, то сила стимулов д.б. наивысшей на экваторе клетки. С некоторыми предположениями относительно природы сигнала, её способа высвобождения и распределения микротрубочек, компьютерное моделирование указывает на то, что стимулы деления д. достигать максимума в экваториальной области (Devore et al., 1989; Harris and Gewalt, 1989).
Вторая гипотеза, названая astral relaxation, утверждает, что астральные лучи (т.e., microtubules) вызывают снижение кортикальной контрактильности зависимым от плотности способом. Согласно этой модели плотность астральных лучей выше вблизи полюсов, чем на экваторе, предполагая сферически симметричные звёздочки в сферических клетках. Это м. приводить к тому, полярные области будут менее контрактильными, чем экваториальные и это различие в контрактильности будет индуцировать образование борозды (Fig. 1 B; Wolpert, 1960). Количественное моделирование подтверждает, что эта модель м. в принципе делать возможным формирование борозды, но указывает на то, что необходима позитивная петля обратной связи во время контрактильности для завершения ingression (White and Borisy, 1983; Yoshigaki, 1999).
Эти две модели исходят из противоположных заключений относительно роли астральных микротрубочек, т.к. отличаются в исходных предположениях о распределении микротрубочек, их длине и пути взаимодействия их с кортексом клетки. Сегодня очевидно, что существуют активности, которые связывают микротрубочки от противоположных звездочек и генерируют структуру, которая называется центральным веретеном (известна также как spindle midzone). Эволюционно законсервированный centralspindlin. 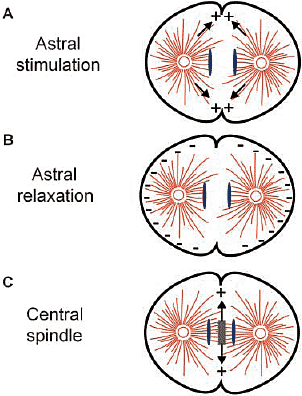 Figure 1. Three schematic models for cleavage furrow positioning. See text for details. комплекс, содержащий kinesin-подобный белок Mklp1 и Rho семейства GAP, HsCYK-4/MgcRacGAP) (Mishima et al., 2002), является одним из таких факторов. Centralspindlin непосредственно участвует в сборке центрального веретена, т.к. он локализуется в центральном веретене и обладает активностью связывания микротрубочек в пучки (Mishima et al., 2002). Др. важным фактором сборки центрального веретена является связывающий микротрубочки белок PRC1 (Mollinari et al.,2002). Имеются доказательства, что антипараллельные пучки микротрубочек м. регулировать формирование борозды, некоторые манипуляции, которые привели к появлению моделей astral stimulation и relaxation д.б. интерпретированы по-новому. В самом деле, наблюдения на Drosophila предоставили доказательства того, что астральные микротрубочки м.б. не критическими для образования борозды и что центральное веретено необходимо и достаточно для индукции образования борозды (Fig. 1 C; Giansanti et al., 2001). В частности, клетки, дефицитные по kinesin-подобному белку Pavarotti (ортологу Hs MKLP1/Ce ZEN-4), не способны формировать центральное веретено, имеют скорее нормальный вид астральных микротрубочек, но не формируют борозды деления (Adams et al., 1998; Somma et al., 2002). Напротив, мутанты asterless, которые лишены большей части астральных микротрубочек, но содержат центральное веретено, всё ещё способны формировать борозду деления (Bonaccorsi et al., 1998). Эти данные не согласуются ни с astral stimulation ни с astral relaxation моделью и подтверждают, что центральное веретено ответственно за индукцию борозды.
Дополнительные доказательства, подтверждают мнение, что центральное веретено участвует в формировании борозды. В культивируемых клетках крыс, если проделываются небольшие перфорации рядом с центральным веретеном, то образование борозды происходит стороне перфорации рядом с центральным веретеном, а не на кортикальном сайте, где д. сформироваться борозда у нормальных клеток (Cao and Wang, 1996). Более того, сперматоциты кузнечиков подвергались манипуляциям по одновременному удалению центросом и хромосом, а оставшиеся микротрубочки само-организуются в пучки, которые напоминают центральное веретено и, по-видимому, индуцируют образование борозды (Alsop and Zhang, 2003). Эти результаты вместе с тем фактом, что многие ключевые регуляторы митотических событий локализуются в центральном веретене, ведут к предположению, что микротрубочки центрального веретена (или более обще, антипараллельные пучки микротрубочек)являются принципиальным регулятором образования борозды.
Однако, имеются убедительные доказательства, что центральное веретено необязательно для образования борозды деления. У эмбрионов Caenorhabditis elegans разрушение центрального веретена не препятствует ingression борозды деления. В этих условиях борозда образуется и сжимается, но она неспособна закончить цитокинез (Powers et al., 1998; Raich et al., 1998; Jantsch-Plunger et al., 2000).
Совершенно др. потребность в центральном веретене для образования борозды у Drosophila и C. elegans из-за различий в размерах клеток в этих системах. Следовательно, критический детерминант для образования борозды м.б. эволюционно не законсервирован. В самом деле, описаны некоторые вариации в локализации критических факторов, регулирующих цитокинез. Напр., у Drosophila
помимо центрального веретена, локализующего пул Pavarotti, имеется также кортикально локализованный пул, которые не выявляется у др. организмов (Sellitto and Kuriyama, 1988; Adams et al., 1998; Powers et al., 1998; Raich et al., 1998; Minestrini et al., 2003). Напротив, в клетках млекопитающих ECT2 (GEF для RhoA) легко обнаруживается в ассоциации и с кортексом клетки и с веретеном, но его ортолог у Drosophila
в первую очередь ассоциирует с кортексом клетки (Prokopenko et al., 1999; Tatsumoto et al., 1999). Однако, недавние результаты указывают на то, что ни размер клеток, ни отсутствие консервации не лежит в основе изменчивой степени, с которой центральное веретено контролирует образование борозды и указывает на то, что этот процесс контролируется двумя параллельными путями. У эмбрионов C. elegans центральное веретено не существенно для образования борозды. Однако, если степень удлинения веретена во время анафазы снижена за счёт некоторых генетических пертурбаций, то центральное веретено становится существенным (Dechant and Glotzer, 2003). Кроме того, хотя образование борозды м. осуществляться в отсутствии центрального веретена, но инициация цитокинеза слегка задерживается в этих условиях. Т.о., вообще-то разные типы клеток используют как астральные микротрубочки, так и центральное веретено для образования борозды, правда, в различной степени. Имеются, доказательства пластичности в индукции бороозды деления у млекопитающих. Показано, что центральное веретено обладает активностью, индуцирующей борозду, даже если клетки истощены по ключевым компонентам центрального веретена, таким как MKLP1 или PRC1, до тех пор, пока формируется борозда (Cao and Wang, 1996; Matuliene and Kuriyama, 2002; Mollinari et al., 2002). Molecular control of furrow formation Принимая во внимание, что и центральное веретено и астральные микротрубочки м. вносить вклад в индукцию борозды деления, по крайней мере, при некоторых условиях, то белки, которые локализуются на этих структурах д.б. потенциальными сигналами для механизма индукции борозды. Центральное веретено в частности содержит многочисленные факторы, участвующие в цитокинезе. В принципе, эти факторы м. регулировать образование борозды двумя способами: они м.б. позитивными индукторами образования борозды или они б. ингибировать негативные регуляторы образования борозды. Delivery of an activator of furrow formation Имеется несколько факторов, которые концентрируются на центральном веретене, которые, как полагают, являются индукторами образования борозды деления. Одним из кандидатов является комплекс ABI, состоящий из Aurora B, INCENP и Survivin/BIR-1 (Adams et al., 2000; Kaitna et al., 2000; Kang et al., 2001; Bolton et al., 2002; Cheeseman et al., 2002; Honda et al., 2003; Romano et al.,2003). У нематод этот комплекс содержит 4 белка, CSC-1 (Romano et al., 2003). В клетках млекопитающих, INCENP сначала локализetncz на хромосомах во время прометафазы, затем он концентрируется на центромерах во время метафазы, а затем в начале анафазы он локализуется и в центральном веретене и, что интересно, в лежащем поверх клеточном кортексе (Cooke et al.,
1987). Как астральные микротрубочки, так и центральное веретено вносят вклад в кортикльную локализацию Aurora B (Murata-Hori and Wang, 2002), преимущественно благодаря взаимодействиям с INCENP и Survivin, чья сольная функция, по-видимому, проявляется в активации и локализации Aurora B. Интересно, что Aurora B локализуется в центральном веретене в клетках, которые лишены хромосом (Bucciarelli et al., 2003), это указыает на то, что эти субклеточные направляющие события независимы. Кортикальная локализация комплекса ABI предшествует ранним стадиям цитокинеза (Eckley et al., 1997). Хотя такая локализация комплекса ABI указывает на то, что он м. управлять образованием борозды деления, но клетки, дефицитные по Aurora B (из-за мутации, RNAi-обусловленной деплеции или химического ингибирования) остаются компетентными к образованию борозды деления (Schumacher et al., 1998; Fraser et al., 1999; Kaitna et al., 2000; Hauf et al., 2003). Следовательно, Aurora B не является обязательной для образования борозды деления. Однако, функциональное перекрываение м. маскировать роль Aurora B в образовании борозды.
Вторым потенциальным активатором образования борозды деления, которые м. связывать центральное веретено с образованием борозды деления является RhoGEF, Pebble. Pebble , как недавно было показано, ассоциирует с Drosophila centralspindlin (Somers and Saint, 2003). Pebble (Hs ECT2/Ce LET-21) является существенным для образования борозды деления, т.к. он является критическим активатором RhoA в цитокинезе (Prokopenko et al., 1999; Tatsumoto et al., 1999). Двугибидный анализ показал, что NH2 окончание Pebble соединяется с NH2-терминальной областью ортолога мух CYK-4, RacGAP50C. Концентрация centralspindlin в midzone веретена м. тем самым рекрутировать Pebble и индуцировать локальную активацию RhoA, которая сопровождается полимеризацией актина и образованием борозды деления. Если это действительно так, то клетки, дефектные по образованию центрального веретена, также д.б. дефектными по образованию борозды. Хотя купирование этих двух процессов наблюдается у Drosophila оно не имеет места у эмбрионов C. elegans или в клетках млекопитающих. Более того, избыточная экспрессия NH2-терминального домена ортолога Pebble, ECT2, вызывает поздние дефекты цитокинеза (Tatsumoto
et al., 1999), и не ожидаются ранние дефекты, если ассоциация Pebble с centralspindlin существенна для пространственной регуляции функции Pebble. Т.о., хотя Pebble и является критическим для образования борозды, его ассоциация с центральным веретеном, по-видимому, не является критической у всех видов. Ассоциация Pebble с centralspindlin м. способствовать продолжающейся ingression борозды деления за счёт поддержания RhoA в активном состоянии. Важно понять взаимодействие между RhoGAP и RhoGEF в этом необычном белковом комплексе.
Local inhibition of a negative regulator
of furrow formation Альтернативным путём регуляции образования борозды является локальное ингибирование негативного регулятора. Эксперименты на клетках млекопитающих и эмбрионах C. elegans указывают на то, что во время цитокинеза микротрубочки ингибируют контрактильность кортекса клетки. Если индуцируемые микротрубочки оказываются необычно короткими из-за продолжительной активации katanin микротрубочки-обслуживающего комплекса, то наблюдаются дополнительные борозды на клеточных полюсах (Fig. 2 A; Kurz et al., 2002). Сходным образом, клетки млекопитающих, формированные к выходу из митоза в отсутствие микротрубочек, подвергаются сильным неорганизованным контракциям (Canman et al., 2000). Эти данные указывают на то, что микротрубочки ингибируют образование борозд. Учитывая, что имеются убедительные доказательства того, что микротрубочки контролируют образование борозд, то вполне возможно, что модуляция их распределения или свойств м. контролировать образование борозд. В принципе связывание микротрубочек в пучки, чтобы сформировать центральное веретено, д.б. количественно и/или качественно регулировать ингибирующие эффекты микротрубочек. Т.о., второй механизм, с помощь которого центральное веретено м. способствовать образованию борозды, м.б. ингибирование этой негативной регуляции. Элонгация веретена и сборка центрального веретена действуют вместе, чтобы создать локальный минимум плотности микротрубочек в позиции клеточного экватора, одинаково удаленным от двух полюсов веретена (Dechant and Glotzer, 2003). Важно, чтобы эти два процесса действовали параллельно, способствуя образованию борозды (Fig. 2 B). Эти наблюдения согласуются с моделью, согласно которой положение борозды деления в экваториальной области предопределяется местом, когда ингибирующий эффект микротрубочек достигает локального минимума (Fig. 2 D). Сборка центрального веретена м. не только влиять на пространственную организацию микротрубочек, она м. также изменять способность микротрубочек ингибировать контрактильность, меняя свойства микротрубочек. Напр., ингибирование кортикальной контрактильности м. влияить на динамику микротрубочек или на ассоциированные с микротрубочками белки. Соединение факторов с концами микротрубочек м. менять их свойства или динамику. В самом деле, ранние работы на клетках млекопитающих показали, что микротрубочки midzone более стабильны, чем в остальной части клетки (Saxton and McIntosh, 1987). В частности, пред ingression борозды деления субнабор микротрубочек вблизи презумптивной борозды существенно более стабильный, чем микротрубочки вблизи полюсов (Fig. 2 C; Canman et al., 2003). Пока неясно, вызывается ли стабилизация микротрубочек в экваториальной области комплексом centralspindlin, но это очень возможно, т.к. онг присутствует здесь, а избыточная экспрессия kinesin субъединицы centralspindlin (MKLP1/Pavarotti) индуцирует гиперстабилизацию микротрубочек (Minestrini et al., 2002). Т.о., соединение centralspindlin с микротрубочками м. индуцировать сборку центрального веретена и одновременно препятствовать зависящей от микротрубочек ингибиции образования борозды.
Механизм, с помощью которого микротрубочки ингибируют кортикальную контрактильность и контролируют цитокинез, неизвестен. RhoA exchange factor, Dm Pebble (Hs ECT2, Ce LET-21) является одной из наиболее высоко стоящих молекул в этом пути и является кандидатом на регуляцию с помощью микротрубочек. Однако, мало известно от том, как этот критический фактор обмена регулируется, за исключением того, что его активность нуждается в фосфорилировании (Tatsumoto et al., 1999). Кроме того, хотя RhoA и некоторые его 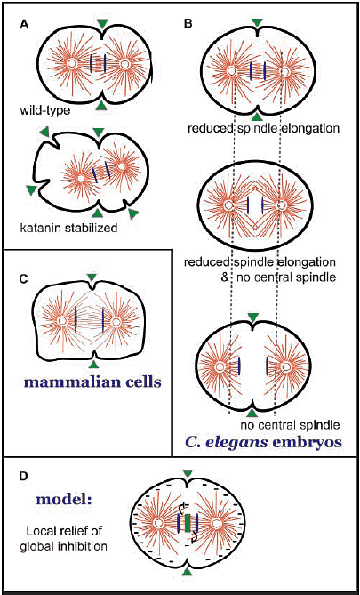 Figure 2. Experimental evidence that furrow positioning may result from local relief from microtubule-dependent inhibition of furrow formation. (A) Stabilization of the microtubule-severing complex katanin leads to microtubule shortening and ectopic furrowing. (B) Either central spindle assembly or spindle elongation are sufficient to induce furrow formation, but when both pathways are inhibited, no furrow formation occurs. (C) In mammalian cells, a subset of the microtubules in the vicinity of the furrow are less dynamic (purple) than elsewhere in the cell. (D) Model for furrow positioning. эффекторы концентрируются в борозде деления, нет прямых доказательств локальных различий в активности RhoA в раннем цитокинезе. Недавно базирующийся на FRET подход позволил наблюдать активный RhoA во время цитокинеза. Он не выявлял обнаружимые количества активного RhoA во время раннего цитокинеза, но активный RhoA появлялся в позднем цитокинезе (Yoshizaki et al., 2003). Т.к. активность RhoA необходима для инициальных стадий образования борозды, то возможно, что низкие уровни активного RhoA управляют образованием борозды и эти уровни RhoA были ниже границы обнаружения в этих зондах. Второе поколение зондов, специфичных к активным формам RhoA и др. молекул, существенных для цитокинеза, м. пролить свет на этот важный вопрос. Однако, учитывая, что имеется несколько прецендентов локальной активации GTPase сигнальных комплексов, то разумно разработать модель, подтверждающую, что локальное увеличение передачи сигналов RhoA индуцирует образование борозды. Если это окажется так, что позиционирование борозды д. объясняться просто, если астральные микротрубочки ингибируют активацию RhoA, ингибируя тем самым образование борозды в эктопических местах, а сборка центрального веретена и элонгация веретена действуют совместно, чтобы обеспечить локальный рельеф этих ингибирующих эффектов и сделать возможной активацию RhoA в экваториальной области. Интересно, что деполимеризация микротрубочек в интерфазных клетках вызывает активацию RhoA-GTP (Ren et al., 1999), указывая тем самым, что микротрубочки м. непосредственно или косвенно ингибировать RhoA. Более того, специальная RhoGEF, GEF-H1, ингибируется с помощью микротрубочками-обуславливаемой секвестрации (Krendel et al., 2002), иллюстрируя один из таких механизмов. Важно, что RhoA м.б. не только фактором, чья активность ограничена пространственно.
Concluding remarks Позиционирование борозды, по-видимому, обусловливается не только индукцией с помощью астральных микротрубочек или центрального веретена, но оба компонента вносят свой вклад. Индукция борозды деления посредством локального содействия со стороны ингибирующих эффектов микротрубочек является привлекательной моделью, т.к. она объясняет как два пути, удлинения веретена и сборки центрального веретена, м. контролировать образование борозды деления посредством общего молекулярного механизма. Кроме того, она объясняет тот факт, что центральное веретено оказывает позитивное (хотя и несущественное) влияние на образование борозды. Она также предсказывает значение локальной деполимеризации микротрубочек для индукции борозды деления.
|