Клетки, которые экспрессируют Shh, представляют собой классическую ЗПА конечностей позвоночных. Многочисленные
эксперименты продемонстрировали, что разные уровни передаваемых сигналов с помощью Shh вдоль передне-задней оси являются критическими для детерминации, какого типа палец будет сформирован. Классически, дифференциальные уровни передачи сигналов, как полагали, устанавливаются с помощью пространственного градиента морфогена. Клетки, подвергшиеся воздействию низких количеств Shh, становятся передними пальцами, тогда как воздействие высоких уровней Shh ведет к образованию задних пальцев. Наши данные указывают, что второй механизм также важен для специализации пальцев с помощью Shh, это временной градиент, устанавливаемый с помощью экспансии производных клеток, экспрессирующих Shh.
В модели временного градиента подразумевается, что клетки внутри ЗПА продолжают обнаруживать высокие уровни передачи сигналов Shh в течение продолжительного периода времени. Некоторые гены, предположительно являющиеся мишенями для Shh, экспрессируются в виде паттерна гнёзд, перекрывающегося с клетками, экспрессирующими Shh в ранних зачатках конечностей, включая
Ptc 1 (Marigo et al., 1996a; Goodrich et al., 1996),
Ptc 2 (Pearse et al., 2001),
Bmp2 (Francis et al., 1994; Laufer et al., 1994) и
Gli (Marigo et al., 1996b). Постепенно один из них,
Gli1, заметно подавляется в области ЗПА (Marigo et al., 1996b). Однако, экспрессия др. предполагаемых мишеней Shh сохраняется в задней части почки конечности с наиболее сильной экспрессией на заднем крае до тех пор, пока экспрессируется
Shh (Marigo et al., 1996a; Pearse et al., 2001; Laufer et al., 1994). Очень возможно, что
Gli1 обнаруживает отличающийся паттерн экспрессии потому, чсто он отвечает только на присутствие активаторов Gli, тjulf как активность др. генов м.б. скорее объяснена отсутствием репрессоров Gli. Позднее в развитии конечности
Gli2 и Gli3 транскрипционно подавляются в области ЗПА (Marigo et al., 1996b). Как следствие этого следует ожидать существенное снижение активности активатора Gli, тогда как отсутствие репрессоров Gli д., пожалуй, укрепляться. Независимо от того, какое отношение
Gli1 имеет к морфогенезу нормальной конечности (Park et al.,2000) функциональное значение его подавления в задней части конечности неясно. Более важна строгая, сохраняющаяся градированная кзади экспрессия
Ptc 1 и др. мишеней, указывающая, что имеется продолжающаяся активная реакция на Shh в области ЗПА в течение всей фазы формирования паттерна конечности.
Fate Map of the ZPA
Зачаток конечности кур очень детально картирован в отношении судеб с помощью Dil (Vargesson et al., 1997). Они установили, что если клетки, которые как ожидалось экспрессируют Shh, были мечены на стадии 20, то м. было видеть, что они расширяются, чтобы образовать домен, скорее всего подобный одному из описанных здесь, на ст. 28. Nj? что задние пальцы целиком происходят из клеток, экспрессирующих Shh, не было оценено по достоинству, однако, благодаря др. экспериментам, показавшим, что первые клетки, экспрессирующие Shh, м. все подвергаться апоптозу в позднем зачатке конечности (Sanz-Ezquerro, Ticle, 2000), указывало на то, что задние клетки, меченные Dil, м. маркировать клетки, не экспрессирующие Shh, в задней части, которые перемешаны с клетками ЗПА. Однако, мы подтвердили более ранние предположения (Saunders, Fallon, 1967), что задний апоптоз не является признаком развития конечностей у мышей. В свете этих находок и принимая во внимание законсервированные механизмы формирования паттерна конечностей у видов, мы предположили, что карта Vargesson м.б. принята за основу и что сходное объяснение клеток, экспрессирующих Shh, приложимо и к конечностям кур. Наблюдаемый апоптоз в задней части конечностей кур, фактически, существует на всех стадиях проксимальнее пластинки кисти (Saunders, Fallon, 1967). Наблюдаемый апоптоз в задней части зачатка конечности цыплят м. иметь отношение к элиминации на поздней стадии передач сигналов Shh в проксимальной части конечности (Sanz-Ezquerro, Tickle, 2000), но мы не м. подтвердить, что это не имеет отношения к экспансии первых клеток ЗПА в дистальной части зачатка.
Cellular Memory of Prior Exposure to Shh
Наша модель базируется на способности клеток отвечать дифференциально и на время экспозиции Shh. Воздействие на мезенхиму зачатков конечностей цыплят высокой концентрацией Shh в течение 10 ч не оказывало влияния на формирование паттерна. однако обработанные клетки сохраняли в памяти это воздействие, это обнаруживалось когда они подвергались воздействию той же самой концентрации в течении более 16 ч. Зачатки пальцев предварительно экспозированные Shh воспринимали судьбу более задних пальцев. чем зачатки пальцев, обрабатываемые параллельно но бех предварительной экспозиции. Эти эксперименты показали, что время экспозиции Shh м. в самом деле предопределять качественные особенности пальцев. Предыдущие исследования (Yang et al., 1997) также предоставляли доказательства, что это имеет место. Кусочки, пропитанные высокими концентрациями Shh? имплантировали в переднюю часть зачатка конечности цыпленка. Клетки, соседние с кусочком были маркированы Dil. Кусочек затем удаляли после разных промежутков времени. В зависимости от продолжительности экпозиции Shh маркированные клетки развивались в эктопический палец 2 ( короткая экспозиция) или палец 3 или 4 (более продолжительная экспозиция). Эти данные подтвердили модель 'promotion' (Tickle, 1995), согласно которой зачатки пальцев сначала специфицируются в судьбу переднего и затем продвигаются к получению судьбы более задних пальцев с увеличением экспозиции. Модель также постулировала, что разные зачатки пальцев воспринимают разной продолжительности экспозиции Shh благодаря экспансии задней части ткани.
Мнение, изложенное здесь, не совпадает с аспектами 'promotion' (Tickle, 1995). Обе модели учитывают время экспозиции, исходя из экспансии поля конечности в качестве ключевого параметра. Различия в том, что модель 'promotion' фокусируется на последовательных ступенях, с помощью которых качественные особенности пальцев специфицируются в мезенхиме конечности и что согласно этой модели, зачатки всех пальцев зависят как от времени, так и концентрации экспозиции. Напротив, наша модель не совсем ясно объясняет являются или нет задние пальцы сначала временно специфицированными как передние. Однако, она более конкретно подтверждает, что два наиболее задних пальца отличаются только продолжительностью времени, в течение которого они экспозируются высокими уровнями Shh? тогда как спецификация пальца 3 зависит исключительно от низких концентраций, достигаемых только в результате градиента пространственной диффузии.
Shh Directly Specifies Digit Identity
Пока неясно, действует ли Shh непосредственно на формирование передне-задней оси или действует посредством вторичного морфогена (чаще всего предполагается секретируемый фактор BMP2, который в самом деле индуцирует более широкий домен в задней части зачатка конечности с помощью Shh (Laufer et al., 1994). Предполагается также, что Shh необходим на длинной дистанции независимым от концентрации образом, чтобы "prime" мезенхиму конечности, тогда как BMP2, индуцируемый высокой концентрацией Shh в задней части зачатка конечности, действует как настоящий градированный сигнал, специфицирующий качественные особенности разных пальцев (Drossopoulou et al., 2000).
Наши генетические эксперименты с понижением уровней диффундирующего Shh белка у мутантных аллелей
Dispatched-1 помимо подтверждения нашей модели временного градиента в задней части почки конечности подтвердили также то, что Shh м.б. непосредственным морфогеном, специфицирующим пальцы. Если бы BMP2 был настоящим морфогеном, то м. было бы ожидать потери пальца 2, т.к. диффузия BMP2 не нарушена у мутантов и , следовательно, порог концентрации, необходимый для спецификации пальца 2, д. достигаться нормально. Для окончательного решения этого вопроса необходимо удаление BMP из зачатка конечности.
Наши эксперименты по снижению доступности белка Shh приводили в результате к специфической потере пальца 2, как и ожидалось согласно модели, что более задние пальцы менее зависимы от диффузии Shh для свой спецификации. Мы пришли к выводу, что палец 2 в частности детерминируется с помощью концентрационного градиента Shh. Зачаток пальца 2 д. действительно отвечать на Shh во время нормального развития конечности. Из Shh нулевого фенотипа становится ясно, что пальцы 2-5 все зависят от активности Shh? но это не обязательно указывает на то. что Shh непосредственно действует на клетки пальца 2. Такие доказательства теперь получены при картировании судеб, аналогичному, описанному здесь, при котором CRE recombinase, управляемая с помощью промотора известной мишени для Shh,
Gli1, была использована для маркирования клеток у репортерных мышей. β-галактозиданое окрашивание конечностей, где CRE был активирован внутри домена Gli1 во время передачи сигналов Shh, показало, что на самом деле пальцы 2-5 возникают из клеток-предшественников, которые активно реагировали на Shh во время развития конечности (Ahn, Joyner, 2004).
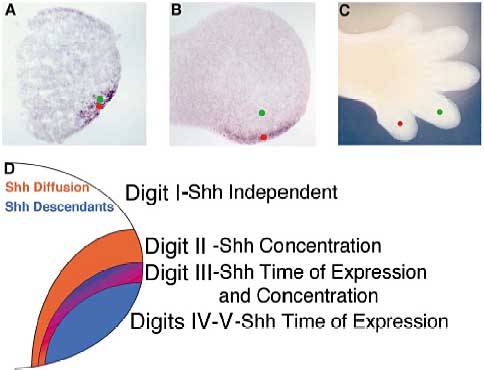 Figure 5. There Is a Temporal Gradient of Shh Exposure in the Posterior Limb Bud
Cells transcribing Shh themselves see the highest levels of Shh signaling in an autocrine manner. Early in limb development, the progenitors of both digit 4 (green dot) and digit 5 (red dot) are within the Shh-expressing ZPA domain (A), since both those digits are identified as Shh descendants in our lineage-tracing experiments. At later stages, as demonstrated by our tamoxifen-activated lineage-tracing experiments,the primordium of digit 5 still expresses Shh, but the primordium of digit 4 does not (B). Thus, the digits form (C) from cells that have seen maximal levels of Shh signaling for different lengths of time (D). Digit 3 contains some cells that have seen maximal Shh signaling for an even shorter period of time than digit 4 but also depends on low levels of Shh signaling from diffusion, since this digit is lost in mice producing only an altered form of Shh (N-Shh), which does not signal outside of the cells that produce it. Digit 2 is completely dependent upon Shh diffusion for its specification, as it is lost in Shh null animals but does not contain any cells descended from the Shh-expressing domain. See
text for details.
Figure 5. There Is a Temporal Gradient of Shh Exposure in the Posterior Limb Bud
Cells transcribing Shh themselves see the highest levels of Shh signaling in an autocrine manner. Early in limb development, the progenitors of both digit 4 (green dot) and digit 5 (red dot) are within the Shh-expressing ZPA domain (A), since both those digits are identified as Shh descendants in our lineage-tracing experiments. At later stages, as demonstrated by our tamoxifen-activated lineage-tracing experiments,the primordium of digit 5 still expresses Shh, but the primordium of digit 4 does not (B). Thus, the digits form (C) from cells that have seen maximal levels of Shh signaling for different lengths of time (D). Digit 3 contains some cells that have seen maximal Shh signaling for an even shorter period of time than digit 4 but also depends on low levels of Shh signaling from diffusion, since this digit is lost in mice producing only an altered form of Shh (N-Shh), which does not signal outside of the cells that produce it. Digit 2 is completely dependent upon Shh diffusion for its specification, as it is lost in Shh null animals but does not contain any cells descended from the Shh-expressing domain. See
text for details.
Temporal Signaling Explains Phenotypes that Are Difficult to Understand on the Basis of a Spatial Morphogenic Gradient
Модель временного градиента, предложенная здесь, предоставляет объяснение аспектов фенотипа др. мутантов, затрагивающих активность Shh в зачатках конечностей. Shh имеет два основных домена: каталитический С-терминальную половину и N-терминальную половину, которая содержит сигнальную активность. Если этот N-терминальный домен экспрессируется в клетках млекопитающих, то он собственно не модифицируется, хотя он сохраняет свою сигнальную потенцию. Если аллель, экспрессирующий только N-терминальную порцию Shh помещается на место нормального аллеля Shh, то хотя и синтезируются нормальные уровни N-Shh, однако он способен только передавать сигнал в клетках, которые его продуцируют (Lewis et al., 2001). Т.о., вместо того, чтобы достигать генов, таких как
Ptc 1, экспрессируемого в широком, градированном домене в задней части почки конечности, они экспрессируются в небольшом резко очерченном заднем домене, идентичном домену самого
Shh. Скелетный фенотип, наблюдаемый у таких животных характеризуется потерей пальцев 2 и 3. Сохранение пальца 1 ожидается, т.к. он независим от Shh, а сохранение пальцев 4 и 5 объясняется тем, что эти два пальца формируются целиком из клеток, экспрессирующих Shh. Однако, т.к. пальцы 4 и 5 имеют разную морфологию, то они возникают в отсутствие функционального транспорта белка Shh, значит дифференциальная спецификация этих двух пальцев не м. базироваться на классическом диффундирующем градиента, необходим др. механизм. Наши данные, продемонстрировавшие различия во времени экспозиции двух зачатков, делают возможным объяснение: у этих мутантов, как у в диком типе, зачатки пальцев 4 и 5 подвергаются действию максимальных уровней Shh в разной продолжительности промежутки времени, и временной градиент экспозиции Shh является ключевым параметром для дискриминации между качественными особенностями этих двух наиболее задних пальцев.
Conclusion
Итак, наши результаты согласуются со спецификацией пальцев 5-2 предопределяемой с помощью дозы Shh, получаемой мезенхимой почки конечности. однако, они указывают на то, что доза критически зависит от времени экспозиции в дополнение к концентрации Shh белка, доступного разным частям почки конечности. В частности, спецификация пальца 2 достигается с помощью пространственного градиента, тогда как дифференциальная спецификация между пальцами 4 и 5 предопределяется разным временим экспозиции, временным градиентом. Палец 3 м.б. специфицирован комбинацией этих двух параметров экспозиции в пространстве и времени.
Сайт создан в системе
uCoz