Notch Signaling Can Regulate Entoderm Formation in Zebrafish Dev.Dyn. V. 229. No. 4, P. 756-762. 2004. | |
|
Энтодерма возникает из наиболее маргинальных бластомеров у эмбрионов на ст. бластулы. Установлено, что на ст. средней бластулы одиночные клетки м. давать и энтодерму и мезодерму. На ст. поздней бластулы одиночные клетки обычно дают или энтодерму или мезодерму, указывая тем самым что клоны разделились. Однако, механизм, ведущий к сегрегации энтодермы и мезодермы неизвестен.
Предшественниками энтодермы являются не более 4-х клеток, удаленных от края бластодермы и перемешанных с клетками, которые дают некоторые мезодермальные производные, такие как сомиты и сердце, указывая тем самым, что энтодермальные и мезодермальные предшественники перемешаны до некоторой степени вдоль края бластодермы.
Генетический анализ у рыбок данио выявил три гена (bonnie and clyde (bon), faust (fau) и casanova (cas), которые играют существенную роль в формировании энтодермы. Мутанты bon и fau содержат 10% и 60% от нормального количества энтодермальных клеток, соотв., тогда как мутантны cas лишены энтодермальных клеток. Гены bon, fau и cas кодируют Mix-типа гомеодоменовый транскрипционный фактор, цинковые пальчики транскрипционный фактор Gata5 и high-mobility group (HMG) доменовый транскрипционный фактор, соотв. Недавно выявлен еще один Mix-типа гомеодоменовый транскрипционный фактор, Mezzo, участвующий в формировании энтодермы, с частично перекрывающейся функцией с Bon. Анализ экспрессии генов и эксперименты по неправильной экспрессии показали, что эти 4 транскрипционных фактора действуют ниже пути передачи сигналов Nodal и выше sox17, гена HMG транскрипционного фактора и гена foxA2 , winged helix/forkhead транскрипционного фактора ( известного также как axial)/ экспрессирующихся в энтодермальных клетках.
Установлено, что передача сигналов Notch играет важную роль в спецификации судеб клеток во время развития. Механизм латеральной ингибиции использует передачу сигналов Notch, как предполагается, для наделения различными клеточными судьбами соседних клеток. У рыбок данио латеральное ингибирование регулирует развитие первичных нейронов и олигодендроцитов, а также выбор между судьбой hypochord и notochord. Гены bon, mezzo и fau/gata5 экспрессируются во всех клетках по краю окружности, тогда как cas, sox17 и foxA2 экспрессируются в разбросанных энтодермальных клетках по маргинальному домену поздней бластулы и ранней гаструлы. Паттерн соль-и-перец для cas, sox17 и foxA2 указывает на то, что энтодермальные клетоки м.б. отобраны с помощью латеральной ингибиции.
В данной работе проверяли, может ли передача сигналов Notch регулировать формирование энтодермы в маргинальном домене. Активация пути передачи сигналов Notch с помощью избыточной экспрессии Notch intracellular domain (NIVD) у эмбрионов рыбок данио приводила к редукции количества клеток, экспрессирующих sox17 и foxA2. Далее было установлено, что экспрессия bon, который функционирует выше sox17 и foxA2, существенно редуцируется при активации передачи сигналов Notch. Полученные данные указывают на то, что передача сигналов Notch м. влиять на принятие решения стать энтодермой, возможно за счёт отделения её от мезодермы. Попытки блокировать передачу сигналов Notch не приводило к увеличению количества энтодермальных клеток. Возможно, что др. механизмы регулируют количества энтодермальных клеток и что блокирование передачи сигналов Notch недостаточно, чтобы их перекрыть. Нужна техника для активации и блокирования сигналов Notch в одиночной клетке во время принятия решения и способ отслуживать судьбу таких одиночных клеток, чтобы окончательно решить вопрос о роли Notch/ Не исключена возможность, что передача сигналов Notch регулирует временной паттерн экспрессии bon и тем самым количество энтодермальных клеток. Экспрессия bon инициируется в средней бластуле и становится не обнаружимой при 60% epiboly. Полученные данные показывают, что преждевременное активирование сигналов Notch ведет к преждевременному снижению экспрессии bon и последующему снижению количества энтодермальных клеток; т.о., эндогенная передача сигналов Notch м. выключать экспрессию bon и тем самым регулировать формирование энтодермы. И как показывает дальнейшее уменьшение энтодермальных клеток у мутантов bon, которым инъецируется NICD, что сигналы Notch также м. регулировать образование энтодермы и др. bon-независимым способом.
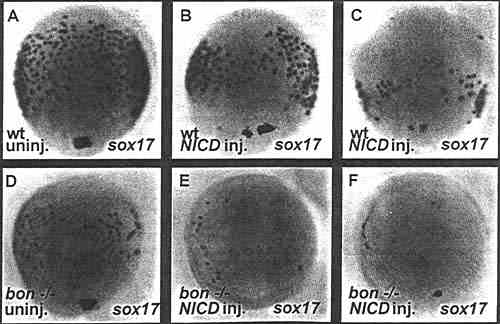 Fig. 4. Notch signaling also functions independently of bon function in endoderm formation. Dorsal views (anterior to the top) of 80% epiboly stage embryos (8.3 hours postfertilization). Embryos are wild-type (wt, A,B,C) or from an incross of bon-/- fish (D.E.F). Embryos were left uninjected (A,D), or injected with 100 pg of NICD mRNA at the one- to four-cell stage (B.C.E.F). A,D: The loss of bon function reduces the number of sox7 7 expressing endodermal cells. B,C: NICD overexpression also reduces the number of sox 17 expressing endodermal cells. E,F: NICD overexpression in bon mutants further reduces the number of sox 17 expressing endodermal cells. | |
)