используется и во время органогенеза позвоночных в тканях, не родственных глазам. В двух таких примерах член семейства Рах генов замещен в такой иерархии. В развивающихся почках гомозиготная делеция
нарушает ранний морфогенез почек (Torres et al., 1996; Xu et al., 1999; Xu et al., 2003). Кроме того, компаундные гетерозиготные мутации Eya1 и Six1 имеют гипопластичные почки (Xu et al., 2003, Li et al., 2003), демонстрируя взаимодействие между этими генами. Т.к. все взаимодействия из
не законсервированы в почках позвоночных, но существует достаточно параллелей для выявления важности
генетической иерархии для органогенеза (Torres et al., 1996; Xu et al., 1999; Xu et al., 2003), здесь
, однако, замещает Pax6. В развивающихся сомитах
действуют синергично, чтобы обеспечить миогенез (Heanue et al., 1999). В этом примере, Pax6 замещается на Pax3 ген.
парадигма используется в органогенезе позвоночных, но иерархия модифицирована за счёт замены соотв. ткане-специфичных членов семейства.
Возможно ли, что non-orthologous
Six гены,
Six3 или Six6, замещают функцию
so таким же образом в глазах позвоночных? Как отмечалось раньше, два
Six гена широко экспрессируются в глазах позвоночных во время морфогенеза,
Six3 и Six6 (Jean et al., 1999). Эти гены, однако, являются ортологами
optix (Kawakami et al., 2000), который экспрессируется во время развития глаз у
Drosophila (Seimiya and Gehring, 2000), но не для
so. Чтобы определить, м. ли
Six3 или Six6 замещать
so в хрусталиках или NR, необходимо знать, ведут ли себя эти Six более сходно с
so или более сходно с
optix во время развития глаз у позвоночных. Для обоих рассматриваемых генов выявлено их качественное и функциональное сходство с
so и с
optix, если учитывать их временной и пространственные паттерны экспрессии, их способность индуцировать эктопические глаза, их гомозиготный нулевой фенотип, их регуляторные взаимоотношения с
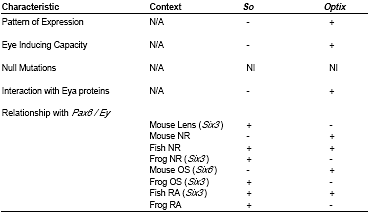
Pax6, и их способность взаимодействовать с белками Eya (Табл. II).
Patterns of expression
Optix и
os имеют разные паттерны экспрессии в развитии глаз
Drosophila (Seimiya and Gehring, 2000).
Optix имеет паттерн экспрессии, сходный с
ey в глазных зачатках и впереди MF в дифференцирующемся диске, тогда как
so обладает паттерном экспрессии, сравнимым с
eya, и обнаруживается в клетках, соседних с и входящих в MF (Seimiya and Gehring, 2000).
Six3 имеет паттерн экспресии почти идентичный таковому у
Pax6, т.к. оба гена обнаруживаются в зачатке сетчатки, LP, и в течение всего развития хрусталикового пузырька и OC (Walther and Gruss, 1991; Oliver et al ., 1995). Экспрессия
Six6 перекрывает экспрессию
Pax6 в производных зачатка сетчатки (optic stalk и neural retina), но отсутствует в действительном зачатке (Toy et al., 1998; Toy and Sundin, 1999; Jean et al., 1999; Bernier et al ., 2000).
Six6 полностью отсутствует в поверхностной эктодерме головы и её производных (Oliver et al., 1995; Toy et al. , 1998; Toy and Sundin, 1999; Jean et al ., 1999).
Six3 и Six6 экспрессируются раньше и более широко, чем
Eya1, Eya2 или Eya3 (Xu et al., 1997). Т.о.,
Six3 и Six6 экспрессируются в виде паттернов, сходных с таковым
Pax6, и , следовательно, в этом отношении более напоминают
optix , чем
so.
Ectopic eye inducing capacity
У мух эктопическая экспрессия одного
optix в антенном диске индуцирует глаза, тогда как эктопическая экспрессия
so этого не вызывает (Seimiya and Gehring, 2000; Pignoni et al., 1997). Подобно
optix, эктопическая экспрессия
Six3 у рыбок medaka индуцирует эктопические примордии сетчатки в компетентных местах внутри головного мозга и со значительно меньшей частотой эктопические хрусталики в головной эктодерме вблизи отических пузырьков (Loosli et
al., 1999). У Xenopus эктопическая экспрессия или
Six3 или Six6
превращает переднюю часть нервной пластинки в сетчатку (Bernier et al., 2000.) В этих экспериментах низкие концентрации Six3 of Six6 расширяют размер сетчатки, тогда как высокие концентрации любого из генов трансформируют средний мозг в сетчатку и удаляют обычные глаза (Bernier et al., 2000). Т.о.,
Six3 и Six6 напоминают
optix по своей способности индуцировать специфичные для глаз программы развития в non-ocular ткани.
Homozygous null phenotypes
Os необходим для всех аспектов развития зрительной системы у
Drosophila (Cheyette et al., 1994; Serikaku and O'Tousa et al. , 1994), тогда как
optix мутантные мыши еще не описаны.
Six6 нулевые мыши имеют гипоплазию сетчатки и у них часто отсутствует зрительный нерв и зрительный перекрест (Li et al., 2002). Кроме того, гетерозиготные мутации
SIX6 у людей коррелируют с anophthalmia (Gallardo et al., 1999). Мутации
SIX3 у людей вызывают holoprosencephaly, с фенотипами, варьирующими от cyclopia до hypotelorism (Wallis et al., 1999; Pasquier et al., 2000). Morpholino ингибирование экспрессии
Six3 у рыбок medaka устраняет и передний мозг и глазную ткань (Carl et al., 2002). Отсутствие
optix у мутантных мух делает неопределенным, напоминают ли мутации
Six3 and Six6 у позвоночных более
so или optix, хотя учитывая тяжесть каждого из описанных нулевых фенотипов мало вероятно, что этот признак м. помочь придти к определенному выводу.
Regulatory relationship to ey/Pax6
У мух экспрессия
optix в само деле не зависит от
ey, , т.к. глаза, эктопически индуцированные с помощью
optix не экспрессируют
ey, и
optix м. индуцировать глаза у
ey дефицитных мух (Seimiya and Gehring, 2000). Напротив, экспрессия
so зависит от
ey (Pignoni et al., 1997; Halder et al., 1998; Niimi et al., 1999; Michaut et al., 2003). Более того,
so действует вместе с
eya, чтобы индуцировать экспрессию
ey в эктопических глазах (Pignoni et al., 1997; Bonini et al., 1997). У позвоночных взаимоотношения
Six3 или Six6 с
Pax6 зависят и от вида и от определённого компартмента развивающихся глах (Table II)
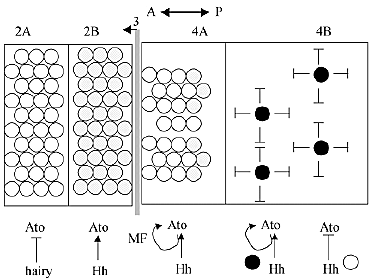 Specification of the R8 founder cell in the Drosophila retina.
Shown is a schematic representation of R8 cell specification in the Drosophila eye disc. The same compartmental nomenclature is used, although
compartments 2 and 4 are expanded into A and B sub compartments to
provide more detail. Anterior (A) is to the left and posterior (P) is to the right. The morphogenetic furrow (MF; striped line at 3 ) progresses anteriorly. Most cells anterior to the MF, in compartment 2A (open circles), do not express ato because it is repressed by hairy. In cells just anterior to the furrow, compartment 2B, Hh induces ato expression. Ato positive cells (dotted circles) are progressively restricted in number, in compartment 4B,until there is one founder cell (filled circles) around which each ommatidia is assembled. The restriction process occurs in four major stages: 1) induction of ato (2B); 2) restriction of ato to an intermediate group of 10-20 cells (not shown); 3 ) restriction of ato to an equivalence group of 2-3 cells (4A); and 4) restriction of ato to a single founder cell, R8 (4B). During R8 differentiation, hh is expressed in cells that do not express ato, and it activates ato in the R8
founder cell and contributes to ato repression in other cells.
Specification of the R8 founder cell in the Drosophila retina.
Shown is a schematic representation of R8 cell specification in the Drosophila eye disc. The same compartmental nomenclature is used, although
compartments 2 and 4 are expanded into A and B sub compartments to
provide more detail. Anterior (A) is to the left and posterior (P) is to the right. The morphogenetic furrow (MF; striped line at 3 ) progresses anteriorly. Most cells anterior to the MF, in compartment 2A (open circles), do not express ato because it is repressed by hairy. In cells just anterior to the furrow, compartment 2B, Hh induces ato expression. Ato positive cells (dotted circles) are progressively restricted in number, in compartment 4B,until there is one founder cell (filled circles) around which each ommatidia is assembled. The restriction process occurs in four major stages: 1) induction of ato (2B); 2) restriction of ato to an intermediate group of 10-20 cells (not shown); 3 ) restriction of ato to an equivalence group of 2-3 cells (4A); and 4) restriction of ato to a single founder cell, R8 (4B). During R8 differentiation, hh is expressed in cells that do not express ato, and it activates ato in the R8
founder cell and contributes to ato repression in other cells.
У мышей
Six6 экспрессируется оптическом стебле и в остатках OV у гомозигот Pax6Sey на E9.5 (Jean et al., 1999), демонстрируя, что
Six6 экспрессия не зависит от
Pax6. Более того,
Six6 нулевые мыши имеют нормальную экспрессию
Pax6 и в хрусталике и сетчатке (Li et al., 2002). Т.к. экспрессия
Six6 и Pax6 независима у мышей, то их взаимоотношения напоминают таковые между
ey и
optix.
Six3 и Pax6, однако, регулируют экспрессию др. др. в производных поверхностной эктодермы у мышей (Goudreau et al., 2002). у Pax6LacZ мышей экспрессия
Six3 не изменена в сетчатке, но снижена в AEL хрусталика (Goudreau et al., 2002). Эктопическая экспрессия или
Pax6 или Six3 в клетках хрусталиковых волокон индуцирует экспрессию др. генов (Goudreau et al., 2002). Т.о., т.к. экспрессия
Six3не зависит от
Pax6 в сетчатке, то взаимоотношения между
Six3 и Pax6 в хрусталиках мышей не зависимы и , следовательно, напоминают таковые
ey-so.
У Xenopus эктопическая экспрессия
Pax6 расширяет экспрессию
Six3 в OV по срединной линии (презумптивный оптический стебель) и снижает экспрессию
Six3 дистальнее (презумптивная NR; Chow et al., 1999). Кроме того, эктопическая экспрессия или
Six3 или Six6 индуцирует эктопические сетчатки, совпадающие с экспансией экспрессии
Pax6 (Bernier et al.,2000). Сходным образом, у рыбок medaka эктопическая экспрессия или
Six3 или Six6 расширяет экспрессию
Pax6 и индуцирует эктопические сетчатки (Loosli et al., 1999). В каждом из этих упомянутых примеров неправильная экспрессия достигалась инъекциями высоких концентраций РНК в бластомеры во время ранней стадии развития. Т.о., экспрессия инъецированного гена не обязательно происходит в физиологической концентрации и не регулируется ни время, ни место экспрессии. Следовательно, такие эксперименты с избыточной экспрессией выявляют взаимоотношения, которые м. появиться в нормальном развитии, но м. и не происходить. Взаимоотношения между
Pax6 и Six3 , предсказываемые с помощью экспериментов избыточной экспрессии подтверждаются, однако, с помощью данных по комплементарно редуцируемой экспрессии. Подавление экспрессии
Six3 с помощью morpholino
вмешивается и в экспрессию
Pax6 , тогда как вмешательство в экспрессию
Pax6 не нарушает экспрессии
Six3 (Carl et al., 2002). Т.о., у Xenopus и рыбок medaka
Pax6 и Six3/Six6 , по крайней мере, частично взаимо-зависят и , следовательно, напоминают
ey и so.
Synergism with eya
Наконец, у Drosophila,
so действует синергично с
eya, как с помощью непосредственного межбелкового взаимодействия, так и кооперативной индукции эктопических глаз (Pignoni et al., 1997).
Optix, однако, не образует межбелкового комплекса с
Eya (Ohto et al., 1999), a ко-экспрессия с
eya не влияет на чатсоту индукции эктопических глаз с помощью
optix (Seimiya and Gehring, 2000).
Six3 также не взаимодействует строго с Eya белками
in vitro (Ohto et al., 1999; Purcell, 2002).
Six6, однако, действует как ко-репрессор с Dach белками и
in vitro и при иммунопреципитации хроматина, указывая тем самым, что это межбелковое взаимодействие происходит и
in vivo (Li et al., 2002). Не выявлено непосредственного взаимодействия между So или Optix и Dac белками у мух. Six-Dach взаимодействие м представлять , следовательно, уникальный для позвоночных признак. Однако, из-за своей неспособности взаимодействовать строго с Eya белками, Six3 и 6 м.б. более сходны с Optix, чем с So.
Т.о., на базе упомянутых выше критериев (суммированных в Табл. II),
Six3 и Six6 в глазах не ведут себя точно так, как
so. Они обладают некоторыми функциональными характеристиками и
optix и so.
В целом, однако, они имеют больше сходства с
optix, чем с
so.
Eyes absent related genes are not critical for vertebrate eye formation
Как установлено выше,
Eya1 и Six3 не взаимодействуют строго на белковом уровне (Ohto et al., 1999; Purcell, 2002). Что же происходит с
eya компонентом
ey парадигмы у позвоночных? Законсервирована ли функция
eya ?
Нарушение активности
eya у мух предупреждает развитие глаз (Bonini et al., 1997), a коллективная экспрессия Eya1, Eya2, и Eya3 охватывает большинство тканей развивающихся глаз мышей. Eya1 экспрессируется в хрусталиках, OS и NR (Xu et al., 1997). Eya2 отсутствует в хрусталиках, но экспрессируется в NR в виде паттерна, комплементарного Eya1 (Xu et al., 1997). В сетчатке Eya1 обнаруживается в ONL и периферических частях сетчатки, тогда как Eya2 обнаруживается в задней области и в INL (Xu et al., 1997). Eya3 присутствует в OV и в периокулярной мезенхиме, но отсутствует в хрусталиках (Xu et al., 1997).
Эти паттерны экспрессии указывают на то, что гены
Eya м. играть роль в морфогенезе глаз позвоночных и что каждый ген
Eya принимает участие в разных компонентах активности глаз
Drosophila. Гомозиготные Eya1 мутанты, однако, имеют только слабый несвойственный глазной фенотип, открытые веки при рождении (Xu et al., 1999), a мыши с компаундными гомозиготными мутациями
Eya1 и Eya2 также имеют морфологически нормальные глаза (P.Y. Xu and R.L. Maas, unpublished; Purcell, 2002). Нулевые мутантные Eya3 мыши еще не получены. Т.о., у мышей Eya1 и Eya2 не являются важными для морфогенеза глаз и , следовательно, не м. играть роль, сопоставимую с соотв. ролью
eya у
Drosophila. Идентифицированы, однако, 3 случая у людей, у которых гетерозиготные мутации
EYA1 коррелировали с дефектами переднего сегмента глаз (Azuma et al., 2000). Дополнительная информация о механизме этого дефекта у людей необходима, т.к. многие др. мутации
EYA1 не обнаруживают влияния на развитие глаз (Vervoot et al., 2002). Возможно, что эти мутации в конечном итоге прольют свет на функцию генов
Eya в глазах позвоночных.
Drosophila eya стоит ниже
ey и является критическим для его функции (Pignoni et al., 1997; Bonini et al., 1997). В самом деле, и частота индукции и размеры эктопических глаз увеличиваются, когда ко-экспрессируются
eya и ey (Bonini et al., 1997). У мышей, ни экспрессия
Eya1, ни
Eya2 не меняется в ранних глазах
Pax6LacZ гетерозиготных мышей (Goudreau et al., 2002). К сожалению, экспрессия генов
Eya позвоночных начинается слишком поздно, чтобы реально испытывать влияние на паттерн экспрессии у гомозиготных
Pax6Sey мышей, т.к. ткани, в которых эти гены экспрессируются, не способны формироваться (Purcell, 2002). Ясно, однако, что если гены
Eya1 и Eya2 м.б. нарушены без влияния на морфогенез глаз (Xu et al ., 1999; Purcell, 2002), значит они не м.б. важными ниже стоящими медиаторами функции
Pax6.
Пока мало доказательств, подтверждающих гомологичные взаимоотношения между генами
Eya позвоночных и
eya мух. Вообще-то наилучшим доказательством того, что эти гены сохраняют некоторую функциональную гомологию, получены в результате демонстрации, что каждый из мышиных генов
Eya1, Eya2 и Eya3 м. восстанавливать формирование глаз у мутантных
eya мух (Bonini et al., 1997; Bui et al., 2000). Мы знаем, однако, что
Eya1 и Eya2 обладают определенной способностью вести себя подобно
Drosophila eya в отновшении
ey парадигмы во время органогенеза в др. тканях позвоночных (Heanue et al., 1999; Xu et al., 1999; Xu et al ., 2003). Тем не менее, способность замещать
eya в глазах
Drosophila не указывает на то, что
Eya1 и Eya2 позвоночных выполняют специфическую роль в развитии глаз позвоночных.
Dach1 is not essential for vertebrate eye formation
У мух паттерн экспрессии
dac совпадает с таковым
eya
(Chen et al., 1997). В глазах позвоночных
Dach1 экспрессируется в хрусталиках и на периферии сетчатки (Caubit et al., 1999; Purcell,2002), сходно с экспрессией
Eya1 (Xu et al., 1997). Т.о.,
Dach1 позвоночных сходен с
dac в отношении их совместной локализации с
Eya1. Экспрессия
Dach1 перекрывается также с экспрессией
Pax6, но существенно задерживается во время его проявления (Hammond et al ., 1998; Caubit et al., 1999; Davis et al., 1999; Heanue et al. , 2002). Однако, т.к. потеря экспрессии
dac нарушает формирование глаз у
Drosophila (Shen and Mardon, 1997; Chen et al., 1997), гомозиготные по мутации
Dach1 не имеют нарушений морфогенеза глаз (Davis et al., 2001). Т.о., в то время как
dac существенен для мух,
Dach1 не обязателен для морфогенеза глаз позвоночных.
У
Drosophila, dac действует ниже
ey, eya и so (Shen and Mardon, 1997; Chen et al., 1997; Michaut et al., 2003). У мышей экспрессия
Dach1 в OC не зависит от
Pax6, т.к. его экспрессия сохраняется в презумптивной NR гомозиготных
Pax6Sey мышей (Heanue et al., 2002). Экспрессия
Dach1, однако. нарушена в эктодерме хрусталиков у гомозиготных
Pax6Sey мышей (Purcell, 2002). Эта потеря экспрессии
Dach1 не является результатом глобальной потери экспрессии генов в покоящейся ткани, т.к. др. гены всё ещё экспрессируются (Purcell, 2002). У мух экспрессия
dac участвует в поддержании экспрессии
ey, eya и so (Chen et al., 1997). У
Dach1 мутантных мышей экспрессия ни
Pax6, ни
Six3 не изменена в развивающихся глазах (Purcell, 2002). Т.о., хотя
Dach1 и стоит ниже
Pax6 в эктодерме хрусталиков, многие аспекты
ey-dac взаимоотношений отсутствуют в глазах позвоночных.
Как и в случае восстановления
Drosophila eya мутантов с помощью
Eya1, 2 и 3, экспрессия
Dach2 у мутантов
dac мух восстанавливает глазной фенотип (Heanue et al., 1999). Это не является неожиданностью, т.к.
Dach2 и его взаимоотношения с
Pax3, Six1, и Eya2 во время сомитогенеза сравнимы с взаимоотношениями в
ey парадигме мух (Heanue et al.,1999). Т.о., способность
Dach2 позвоночных восстанавливать
Drosophila dac глазной фенотип отражает консерватизм
ey парадигмы и в органогенезе позвоночных скорее, чем специфическую консервацию функции
Dach в глазах позвоночных.
Потенциал консервации
ey парадигмы в морфогенезе глаз позвоночных привлекает внимание. Выше мы сравнивали каждый генетический компонент
ey парадигмы с эквивалентными генами позвоночных. Т.к. гомологи всех генов из
ey парадигмы мух экспрессируются во время развития глаз позвоночных, хотя функция каждого из этих генов не строго сохранена. Наиболее наглядным примером отсутствия консервации является неспособность мутаций
Eya1 и Eya2 продуцировать эмбриональный глазной фенотип. Необходимо отметить, однако, что гены позвоночных способны к большинству взаимодействий, имеющих место в глазах
Drosophila. Это указывает на то, что ортологи генов у позвоночных сохраняют свою молекулярную функцию, но что компоненты оказываются , до некоторой степени, не связанными. Кроме того, важно отметить, что некоторые аспекты
ey парадигмы прекрасно законсервированы. В частности
Pax6 очень сильно напоминает
ey, тогда как
Six3 и Six6 обладают некоторыми характеристиками
so. Т.о., несмотря на отсутствие строгой консервации
ey парадигмы, очевидно, что некоторые критические гены-регуляторы глаз присутствуют в морфологически дивергентных глазах мух и позвоночных.
Parallel genetic hierarchies control retinal differentiation in the fly and vertebrates
Интересна др. параллель генетического пути между
Drosophila
и позвоночными, между дифференцировкой R8 фоторецепторов дрозофилы и спецификацией RGC позвоночных. Снова эта консервация проявляется в виде аналогичных структур, которые морфологически совершенно отличны. Ниже мы коротко рассмотрим генетические сходства между путями ретинального развития у
Drosophila и позвоночных. В обеих системах переход от naive, состояния предшественника в proneural состояние контролируется частично с помощью передачи сигналов hedgehog и с помощью антагонистических взаимоотношений между пронейральным транскрипционным активатором и пронейральным репрессором basic helix-loop-helix (bHLH) класса.
В глазном диске
Drosophila MF в пространстве и во времени предшествует волне дифференцировки (Рис. 1,3). Инициация и ход MF зависят частично от передачи сигналов Hh, которые секретируются дифференцирующимися фоторецепторами позади MF (Dominguez and Hafen, 1997; reviewed in Treisman and Heberlein,1998).
Hh инициируют дифференцировку фоторецепторов посредством двух самостоятельных сигналов, дально-действующего и коротко-действующего (Greenwood and Struhl, 1999; Kango-Singh et al., 2003). Decapentaplegic, Dpp, обеспечивает дально-действующий сигнал врнутри MF (Greenwood and Struhl, 1999; Рис. 3) и облегчает сдвиг от naive клеток к pre-proneural (PPN) клеткам. Сдвиг к состоянию PPN маркируется с помощью усиления транскрипции bHLH транскрипционного фактора
hairy (Greenwood and Struhl, 1999; Рис. 3).
Hairy является proneural репрессором и маркирует состояние
PPN (Greenwood and Struhl, 1999). В PPN компартменте клетки выходят из клеточного цикла и готовятся к нейрональной дифференцировке (Greenwood and Struhl, 1999).
Медиатор второго, коротко-действующего сигнала, стоящего ниже Hh, не известен, но использует путь Raf (Greenwood and Struhl, 1999). Результатом этого коротко-действующего сигнала является экспрессия Atonal, bHLH транскрипционного фактора, который индуцирует пронейральное состояние (Jarman et al., 1994).
Hairy и atonal имеют строгую границу экспрессии на границе между PPN и proneural (PN)компартментами (Рис. 3; Greenwood and Struhl, 1999). Клетки, которые не проходят через PPN (
hairy +) к PN (
atonal +) переход не дифференцируются в R8 фоторецепторы (Greenwood and Struhl, 1999). Оказавшись позади MF, экспрессия
atonal постепенно ограничивается из всех клеток в MF одной на кластер, R8 клеткой-основательницей (reviewed in Treisman and Heberlein, 1998; Frankfort and Mardon, 2002). то ограничение частично зависит от передачи сигналов Hh (Dominguez and Hafen, 1997; Greenwood and Struhl, 1999).
A wave of Shh signaling also marks retinal differentiation in vertebrates
У рыбок данио
Shh экспрессируется в ретинальном GCL, вентральнее и назальнее оптического диска (Neumann and Nuesslein-Volhard, 2000). Эта зона
Shh постепенно распространяется поперек сетчатки в виде волны, которая по времени совпадает со спецификацией RGCs (Neumann and Nuesslein-Volhard, 2000). Нарушение экспрессии
Shh когда она расширяется поперек сетчатки блокирует волну дифференцировки RGC, так и продолжающуюся экспрессию
Shh (Neumann and Nuesslein-Volhard,2000). Одинаково с секрецией Hh дифференцирующимися фоторецепторами позади MF у мух, Shh обычно секретируется RGCs
in vivo и это управляет волной дифференцировки поперек сетчатки (Neumann and Nuesslein-Volhard, 2000). Во время волны спецификации RGC количество дифференцирующихся RGCs м. увеличиваться или снижаться при изменении уровней Shh (Zhang and Yang, 2001). Низкие концентрации Shh индуцируют увеличение количества дифференцирующихся RGCs, тогда как высокие концентрации Shh ингибируют дифференцировку RGC и снижают их количества (Zhang and Yang, 2001). Это напоминает дифференцировку фоторецепторов у
Drosophila, где Hh индуцирует экспрессию маркеров R8 нейрональных предшественников, atonal, во всех клетках MF, но мешает экспрессии
atonal в клетках, не выбранных в качестве R8 нейрональных предшественников позади MF (Рис. 3).
Изменения концентрации Hh у
Drosophila меняют количества R8 нейрональных предшественников (Dominguez and Hafen, 1997). Т.о., волна экспрессии и Hh и Shh управляет дифференцировкой, а их концентрации являются критическими для спецификации соотв. количеств или R8 основательниц или RGCs, соотв.
У мышей полное удаление экспрессии
Shh продуцирует один зачаток сетчатки, который не способен подразделяться на симметричные зачатки сетчатки и дает в результате образование одого центрально расположенного оптического бокала (OC) (Chiang, et al., 1996). В зависимости от времени и тяжести фенотипа, вызываемого мутациями Shh у мышей роль Shh в спецификации сетчатки осложнена. Однако, одиночный OC у
Shh мутантных мышей вызывает тяжелую дизморфию: двухслойная структура несоответственно формирует паттерн вдоль проксимо-дистальной оси (Chiang et al., 1996). Специфически NR теряется за счёт RPE, указывая тем самым, что Shh м. играть важную роль в спецификации NR. Кроме того, экспрессия
Shh наблюдается в NR мышей во время спецификации RGC (Jensen and Wallace, 1997). Для получения четких доказательств роли Shh в спецификации сетчатки необходимо получение условных нокаутов.
A family of bHLH genes specifies neuronal identity
Маркер RGC мышей, Math5, является ортологом маркера R8 клеток-предшественников мух, atonal (Brown et al., 1998). Экспрессия
Math5 предшествует экспрессии др. bHLH PN генов (
Ngn2, NeuroD и
Mash1) у мышей (Brown et al., 1998).
Math5 экспрессируется первоначально в сетчатке мышей E11.5 в центральном OC и коррелирует с появлением ранних нейронов (Brown et al., 2001; Wang et al. , 2001). Экспрессия
Math5 распространяется на всю сетчатку в виде волны с пиком на E13.5 (Brown et al., 2001;Wang et al., 2001). Во время появления
RGC на E15.5,
Math5 экспрессируется на периферии сетчатки, где происходит нейрональная спецификация (Brown et al., 2001; Wang et al., 2001). Сходным образом ортологи у рыбок данио и Xenopus гена
atonal, ath5 и Xath5 соотв., предопределяют паттерн нейрональной дифференцировки с помощью своей экспрессии (Kaneker et al., 1997;
Masai et al., 2000). Т.о., паттерны динамической экспрессии пронейральных генов позвоночных
Math5/ath5/Xath5 напоминают экспрессию
atonal, которая перемещается поперек глазного диска мух во фронте
MF, предшествуя дифференцировке фоторецепторов.
Мыши, несущие гомозиготную делецию Math5 теряют свыше 80% клеток, экспрессирующих RGC маркеры. Это ведёт к избытку клеток предшественников в пролиферирующем слое сетчатки (Brown et al., 2001; Wang et al., 2001). Наличие такого избытка клеток ретинальных предшественников после первой волны нейрогенеза ведет к спецификации больших количеств амакринных клеток (Brown et al., 2001; Wang et al., 2001). У рыбок данио мутация
ath5 (lak) устраняет первую волну дифференцировки в сетчатке, которая продуцирует RGCs (Kay et al., 2001). Сходным образом клетки в глазном диске
Drosophila, которые не совершают перехода transition к
ato+ состоянию, не м. дифференцироваться в фоторецепторы (Greenwood and Struhl, 1999).
В развивающейся сетчатке позвоночных нейрональный репрессор
Hes1, гомолог мышиного репрессора
hairy, экспрессируется в вентрикулярной зоне и отсутствует в GCL (Tomita et al., 1996).
Hes1 позитивные клетки остаются в пролиферативном слое и не дифференцируются с нейроны (Tomita et al., 1996). У
Hes1 дефицитных мышей волны дифференцировки, которые продуцируют разные нейроны сетчатки, существенно ускоряются (Tomita et al., 1996) и нейрогенез осуществляется за счет пролиферации в RPCs. Потеря обоих пронейральных репрессорных белков (
Hairy и Extramacrochaete) у
Drosophila также ведет к преждевременной дифференцировке фоторецепторов (Brown et al., 1995). У
Hes1-/- мышей меньшее количество клеток предшественников доступно для спецификации на каждой стадии, так что возникающая в результате сетчатка имеет значительно меньше нейронов (Tomita et al., 1996). Исходя из этих данных Hes1, как полагают, репрессирует нейрогенез в пролиферирующих клетках, тем самым предупреждает преждевременную дифференцировку. В согласии с этой идеей находится и избыточная экспрессия
Hes1 в предшественниках сетчатки у постнатальных крыс, которая увеличвает количество Мёллеровой глии за счёт нейронов (Furukawa et al., 2000).
Hes1 и Math1 обнаруживают антагонистические взаимоотношения в нейрогенезе сетчатки позвоночных.
Hes1 экспрессируется в клетках предшественниках и репрессирует преждевременную дифференцировку, тогда как
Math5 экспрессируется в ранних нейрональных предшественниках и способствует дифференцировке RGC. Кроме того, у
Pax6Sey мышей присутствует меньше Math5 экспрессирующих клеток, тогда как домен экспрессии
Hes1 расширяется (Brown et al., 1998). У гомозиготных
Pax6Sey мышей экспрессия
Math5 отсутствует (Brown et al., 1998). Т.к. потеря экспрессии
Math5 коррелирует с экспансией
Hes1, то эти данные предоставляют дополнительные доказательства антагонистических взаимоотношений между этими генами у позвоночных. Эти антагонистические взаимоотношения сильно напоминают таковые между
hairy и atonal у мух.
Summary
In conclusion, we have reviewed the ey paradigm as characterized
in Drosophila, and we have evaluated its potential conservation
in vertebrates. The evidence to date does not support the idea
that the entire ey genetic hierarchy is conserved in the vertebrate
eye. On the other hand, some genetic parallels do exist, as Pax6
activity is highly reminiscent of
ey, and
Six3 and Six6 have some characteristics of so. Nonetheless, it is the overall epistatic relationship amongst the vertebrate homologues of
ey, so, eya, and dac that appears to be specifically absent in the vertebrate eye.
We have also briefly reviewed some aspects of neuronal specification
in the vertebrate retina, as the control of both cell proliferation
and RGC identity by bHLH transcription factors is highly
reminiscent of R8 photoreceptor differentiation in the fly. Thus in
both cases, specific features of developmental regulatory cassettes
have been retained and re-deployed in vertebrate ocular
organogenesis.
As vertebrate geneticists, how then do we best utilize the wealth
of genetic information that is available to us? The aim is to continue
to utilize the wealth of knowledge emerging from studies in Drosophila
to guide our inquiries into mechanisms of vertebrate development.
In the event that a genetic cassette is not maintained, this too
is instructive, as it indicates that the particular role that the genetic
hierarchy evolved to accomplish is either not relevant, or not
sufficient, to meet the complexity of the vertebrate system. It is,
however, increasingly clear that genetic pathways orchestrating
Drosophila eye formation have been adapted in vertebrates for multiple organogenic processes, and that they have been partially
maintained in the eye, despite the significant divergence of vertebrate
and invertebrate eyes.
Сайт создан в системе
uCoz 
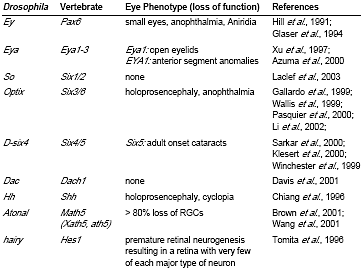 DROSOPHILA EYE SPECIFICATION GENES AND THEIR VERTEBRATE COUNTERPARTS
DROSOPHILA EYE SPECIFICATION GENES AND THEIR VERTEBRATE COUNTERPARTS 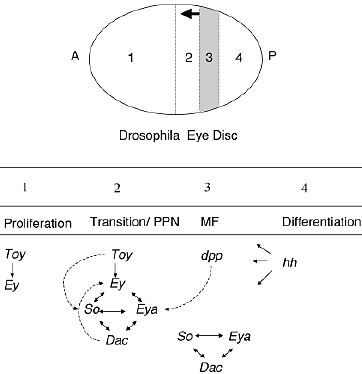 Fig. 1. Schematic representation of Drosophila eye development. Differentiation of the Drosophila eye is controlled by a complex series of signaling events that produce precise compartmentalization of transcription factor activity. The MF, marked by hatched lines, is a wave of differentiation that moves from posterior (P) towards anterior (A) across the eye field during the third instar larvae. Compartments of the eye disc are divided by a dotted line. Compartment 1 represents the majority of cells anterior to the MF. The pre-proneural (PPN) region, represented by compartment 2, is just anterior to the MF. Compartment 3 represents the MF. The arrow indicates the direction of furrow progression. In the compartment 4, posterior to the MF, photoreceptor differentiation and ommatidia assembly occur. Below both
the major cellular events and the expression domains of ey paradigm genes
are indicated. Ey and toy are only expressed anterior to the MF. In the PPN
region, ey and toy induce expression of so, while ey and dpp induce
expression of eya. Together ey, so, and eya activate dac. The nonlinear
regulatory relationship amongst these genes is illustrated and is herein
referred to as the eyeless paradigm. Just posterior to the MF furrow so, eya, and dac continue to be expressed in the absence of eye.
Fig. 1. Schematic representation of Drosophila eye development. Differentiation of the Drosophila eye is controlled by a complex series of signaling events that produce precise compartmentalization of transcription factor activity. The MF, marked by hatched lines, is a wave of differentiation that moves from posterior (P) towards anterior (A) across the eye field during the third instar larvae. Compartments of the eye disc are divided by a dotted line. Compartment 1 represents the majority of cells anterior to the MF. The pre-proneural (PPN) region, represented by compartment 2, is just anterior to the MF. Compartment 3 represents the MF. The arrow indicates the direction of furrow progression. In the compartment 4, posterior to the MF, photoreceptor differentiation and ommatidia assembly occur. Below both
the major cellular events and the expression domains of ey paradigm genes
are indicated. Ey and toy are only expressed anterior to the MF. In the PPN
region, ey and toy induce expression of so, while ey and dpp induce
expression of eya. Together ey, so, and eya activate dac. The nonlinear
regulatory relationship amongst these genes is illustrated and is herein
referred to as the eyeless paradigm. Just posterior to the MF furrow so, eya, and dac continue to be expressed in the absence of eye. 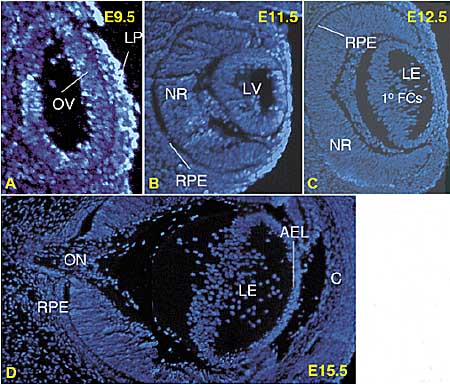 Fig. 2. Vertebrate eye formation. Four key stages of embryonic mouse eye development are shown. (A E9.5; B E11.5; C E12.5; and D E15.5). Each panel shows a representative DAPI stained section through the eye of a paraffin embedded embryo. The vertebrate eye is formed from two separate tissues the neurectoderm and the head surface ectoderm. The retinal anlage, specified in the anterior neurectoderm, is divided into two distinct fields (not shown). From each field an optic vesicle
(OV) evaginates laterally and opposes the overlying surface ectoderm
(SE) (A). In the mouse, the surface ectoderm is induced to form the
lens placode (LP) at roughly E9. The OV is patterned proximal-distally
into optic stalk (OS) and optic cup (OC, not shown), which is subsequently
divided into retinal pigmented epithelium (RPE) and neural retina (NR) (B). The OS matures to form the optic nerve (ON, D). The LP invaginates and forms a hollow lens vesicle (LV; panel B), which is subsequently filled with differentiating primary fiber cells (1o FC) that
elongate from the posterior (C). In the mature lens (LE), an anterior
layer of proliferative epithelial cells (APE) remains and the remainder
of the lens is composed of fiber cells (D). The surface ectoderm from
which the LV pinches off from gives rise to the cornea (C in D).
Fig. 2. Vertebrate eye formation. Four key stages of embryonic mouse eye development are shown. (A E9.5; B E11.5; C E12.5; and D E15.5). Each panel shows a representative DAPI stained section through the eye of a paraffin embedded embryo. The vertebrate eye is formed from two separate tissues the neurectoderm and the head surface ectoderm. The retinal anlage, specified in the anterior neurectoderm, is divided into two distinct fields (not shown). From each field an optic vesicle
(OV) evaginates laterally and opposes the overlying surface ectoderm
(SE) (A). In the mouse, the surface ectoderm is induced to form the
lens placode (LP) at roughly E9. The OV is patterned proximal-distally
into optic stalk (OS) and optic cup (OC, not shown), which is subsequently
divided into retinal pigmented epithelium (RPE) and neural retina (NR) (B). The OS matures to form the optic nerve (ON, D). The LP invaginates and forms a hollow lens vesicle (LV; panel B), which is subsequently filled with differentiating primary fiber cells (1o FC) that
elongate from the posterior (C). In the mature lens (LE), an anterior
layer of proliferative epithelial cells (APE) remains and the remainder
of the lens is composed of fiber cells (D). The surface ectoderm from
which the LV pinches off from gives rise to the cornea (C in D).
 QUALITATIVE AND FUNCTIONAL COMPARISON OF SIX3 AND SIX6 TO SO AND OPTIX
QUALITATIVE AND FUNCTIONAL COMPARISON OF SIX3 AND SIX6 TO SO AND OPTIX