Gene Expression in the Normal Adult Human Kidney Assessed by Complementary DNA Microarray
Mol.Biol.Cell 2004 Feb; 15(2): 649-656 | |
|
The kidney is a highly specialized organ with a complex, stereotyped architecture and a great diversity of functions and cell types. Because the microscopic organization of the nephron, the functional unit of the kidney, has a consistent relationship to the macroscopic anatomy of the kidney, knowledge of the characteristic patterns of gene expression in different compartments of the kidney could provide insight into the functions and functional organization of the normal nephron. We studied gene expression in dissected renal lobes of five adult human kidney using cDNA microarrays represented approximately 30 000 different human genes. Total RNA was isolated from sections of inner and outer cortex, inner and outer medulla, papillary tips, and renal pelvis and from glomeruli isolated by sieving. The results revealed unique and highly distinctive patterns of gene expression for glomeruli, cortex, medulla, papillary tips, and pelvic samples. Immunohistochemical staining using selected antisera confirmed differential expression of several cognate proteins and provided histological localization of expression within the nephron. The distinctive patterns of gene expression in discrete portions of kidney may serve as a resource for further understanding of renal physiology and the molecular and cellular organization of the nephron.
REFERENCES Akaiwa, M., Yae, Y., Sugimoto, R., Suzuki, S.O., Iwaki, T., Izuhara, K., and Hamasaki, N. (1999). Hakata antigen, a new member of the ficolin/opsonin p35 family, is a novel human lectin secreted into bronchus/alveolus and bile. J Histochem Cytochem 47, 777-786. Alizadeh, A.A., et al. (2000). Distinct types of diffuse large B-cell lymphoma identified by gene expression profiling. Nature 403, 503-511. Amara, N., Palapattu, G.S., Schrage, M., Gu, Z., Thomas, G.V., Dorey, F., Said, J., and Reiter, R.E. (2001). Prostate stem cell antigen is overexpressed in human transitional cell carcinoma. Cancer Res 61, 4660-4665. Arystarkhova, E., Wetzel, R.K., Asinovski, N.K., and Sweadner, K.J. (1999). The gamma subunit modulates Na(+) and K(+) affinity of the renal Na,K-ATPase. J Biol Chem 274,33183-33185. Bonilha, V.L., and Rodriguez-Boulan, E. (2001). Polarity and developmental regulation of two PDZ proteins in the retinal pigment epithelium. Invest Ophthalmol Vis Sci 42, 3274-3282. Bonthron, D.T., Brady, N., Donaldson, I.A., and Steinmann, B. (1994). Molecular basis of essential fructosuria: molecular cloning and mutational analysis of human ketohexokinase (fructokinase). Hum Mol Genet 3, 1627-1631. Buhler, R., Pestalozzi, D., Hess, M., and Von Wartburg, J.P. (1983). Immunohistochemical localization of alcohol dehydrogenase in human kidney, endocrine organs and brain. Pharmacol Biochem Behav 18 Suppl 1, 55-59. Canessa, C.M., Schild, L., Buell, G., Thorens, B., Gautschi, I., Horisberger, J.D., and Rossier, B.C. (1994). Amiloride-sensitive epithelial Na+ channel is made of three homologous subunits. Nature 367, 463-467. Cao, Y., Karsten, U., Zerban, H., and Bannasch, P. (2000). Expression of MUC1, Thomsen-Friedenreich-related antigens, and cytokeratin 19 in human renal cell carcinomas and tubular clear cell lesions. Virchows Arch 436, 119-126. Chu, F.F., Doroshow, J.H., and Esworthy, R.S. (1993). Expression, characterization, and tissue distribution of a new cellular selenium-dependent glutathione peroxidase, GSHPx- GI. J Biol Chem 268, 2571-2576. Cox, T.M., Camilleri, M., O'Donnell, M.W., and Chadwick, V.S. (1982). Pseudodominant transmission of fructose intolerance in an adult and three offspring: Heterozygote detection by intestinal biopsy. N Engl J Med 307, 537-540. Danpure, C.J., and Jennings, P.R. (1986). Peroxisomal alanine:glyoxylate aminotransferase deficiency in primary hyperoxaluria type I. FEBS Lett 201, 20-24. Dumont, D.J., Anderson, L., Breitman, M.L., and Duncan, A.M. (1994). Assignment of the endothelial-specific protein receptor tyrosine kinase gene (TEK) to human chromosome 9p21. Genomics 23, 512-513. Eisen, M.B., Spellman, P.T., Brown, P.O., and Botstein, D. (1998). Cluster analysis and display of genome-wide expression patterns. Proc Natl Acad Sci U S A 95, 14863-14868. Floege, J., Alpers, C.E., Sage, E.H., Pritzl, P., Gordon, K., Johnson, R.J., and Couser, W.G. (1992). Markers of complement-dependent and complement-independent glomerular visceral epithelial cell injury in vivo. Expression of antiadhesive proteins and cytoskeletal changes. Lab Invest 67, 486-497. Fukutomi, T., Ando, B., Sakamoto, S., Sakai, H., and Nawata, H. (1996). Thermolabile beta-2 macroglycoprotein (Hakata antigen) in liver disease: biochemical and immunohistochemical study. Clin Chim Acta 255, 93-106. Garber, M.E., et al. (2001). Diversity of gene expression in adenocarcinoma of the lung. Proc Natl Acad Sci U S A 98, 13784-13789. Gollub, J., et al. (2003). The Stanford Microarray Database: data access and quality assessment tools. Nucleic Acids Res 31, 94-96. Hata, A., Lagna, G., Massague, J., and Hemmati-Brivanlou, A. (1998). Smad6 inhibits BMP/Smad1 signaling by specifically competing with the Smad4 tumor suppressor. Genes Dev 12, 186-197. Higgins, J.P., Shinghal, R., Gill, H., Reese, J.H., Terris, M., Cohen, R.J., Fero, M., Pollack, J.R., Van De Rijn, M., and Brooks, J.D. (2003). Gene expression patterns in renal cell carcinoma assessed by complementary DNA microarray. Am J Pathol 162, 925-932. Higgins, J.P., and Warnke, R.A. (1999). CD30 expression is common in mediastinal large B-cell lymphoma. Am J Clin Pathol 112, 241-247. Inaba, S., Okochi, K., Yae, Y., Niklasson, F., and de Verder, C.H. (1990). Serological studies of an SLE-associated antigen-antibody system discovered as a precipitation reaction in agarose gel: the HAKATA antigen-antibody system. Fukuoka Igaku Zasshi 81, 284-291. Kaplan, J.M., et al. (2000). Mutations in ACTN4, encoding alpha-actinin-4, cause familial focal segmental glomerulosclerosis. Nat Genet 24, 251-256. Koszdin, K.L., and Bowen, B.R. (1992). The cloning and expression of a human alpha-1,3 fucosyltransferase capable of forming the E-selectin ligand. Biochem Biophys Res Commun 187, 152-157. Kullak-Ublick, G.A., Hagenbuch, B., Stieger, B., Schteingart, C.D., Hofmann, A.F., Wolkoff, A.W., and Meier, P.J. (1995). Molecular and functional characterization of an organic anion transporting polypeptide cloned from human liver. Gastroenterology 109, 1274-1282. Kurihara, H., Anderson, J.M., Kerjaschki, D., and Farquhar, M.G. (1992). The altered glomerular filtration slits seen in puromycin aminonucleoside nephrosis and protamine sulfate-treated rats contain the tight junction protein ZO-1. Am J Pathol 141, 805-816. Lemley, K.V and Kriz, W. (1991). Anatomy of the renal interstitium. Kidney Int 39, 370-381. Liu, Y., et al. (2000). Edg-1, the G protein-coupled receptor for sphingosine-1-phosphate,is essential for vascular maturation. J Clin Invest 106, 951-961. Lobban, E.D., Smith, B.A., Hall, G.D., Harnden, P., Roberts, P., Selby, P.J., Trejdosiewicz, L.K., and Southgate, J. (1998). Uroplakin gene expression by normal and neoplastic human urothelium. Am J Pathol 153, 1957-1967. Lu, P.C., Ye, H., Maeda, M., and Azar, D.T. (1999). Immunolocalization and gene expression of matrilysin during corneal wound healing. Invest Ophthalmol Vis Sci 40, 20-27. Mannervik, B. (1985). The isoenzymes of glutathione transferase. Adv Enzymol Relat Areas Mol Biol 57, 357-417. Matsumura, Y., et al. (1999). Overt nephrogenic diabetes insipidus in mice lacking the CLC-K1 chloride channel. Nat Genet 21, 95-98. Misra, R.P. (1972). Isolation of glomeruli from mammalian kidneys by graded sieving. Am J Clin Pathol 58, 135-139. Nath, R., Kambadur, R., Gulati, S., Paliwal, V.K., and Sharma, M. (1988). Molecular aspects, physiological function, and clinical significance of metallothioneins. Crit Rev Food Sci Nutr 27, 41-85. Nichols, B.L., Avery, S., Sen, P., Swallow, D.M., Hahn, D., and Sterchi, E. (2003). The maltase-glucoamylase gene: common ancestry to sucrase-isomaltase with complementary starch digestion activities. Proc Natl Acad Sci U S A 100, 1432-1437. Perou, C.M., et al. (2000). Molecular portraits of human breast tumours. Nature 406, 747-752. Radominska-Pandya, A., Little, J.M., and Czernik, P.J. (2001). Human UDPglucuronosyltransferase 2B7. Curr Drug Metab 2, 283-298. Reczek, D., Berryman, M., and Bretscher, A. (1997). Identification of EBP50: A PDZcontaining phosphoprotein that associates with members of the ezrin-radixin-moesin family. J Cell Biol 139, 169-179. Rodriguez-Barbero, A., Obreo, J., Eleno, N., Rodriguez-Pena, A., Duwel, A., Jerkic, M., Sanchez-Rodriguez, A., Bernabeu, C., and Lopez-Novoa, J.M. (2001). Endoglin expression in human and rat mesangial cells and its upregulation by TGF-beta1. Biochem Biophys Res Commun 282, 142-147. Ross, D.T., et al. (2000). Systematic variation in gene expression patterns in human cancer cell lines. Nat Genet 24, 227-235. Roy-Chaudhury, P., Simpson, J.G., and Power, D.A. (1997). Endoglin, a transforming growth factor-beta-binding protein, is upregulated in chronic progressive renal disease. Exp Nephrol 5, 55-60. Schena, M., Shalon, D., Davis, R.W., and Brown, P.O. (1995). Quantitative monitoring of gene expression patterns with a complementary DNA microarray. Science 270, 467-470. Schnabel, E., Anderson, J.M., and Farquhar, M.G. (1990). The tight junction protein ZO-1 is concentrated along slit diaphragms of the glomerular epithelium. J Cell Biol 111, 1255-1263. Simon, D.B., Karet, F.E., Hamdan, J.M., DiPietro, A., Sanjad, S.A., and Lifton, R.P. (1996). Bartter's syndrome, hypokalaemic alkalosis with hypercalciuria, is caused by mutations in the Na-K-2Cl cotransporter NKCC2. Nat Genet 13, 183-188. Stolz, A., Hammond, L., Lou, H., Takikawa, H., Ronk, M., and Shively, J.E. (1993). cDNA cloning and expression of the human hepatic bile acid-binding protein. A member of the monomeric reductase gene family. J Biol Chem 268, 10448-10457. Suliman, Y., Opitz, O.G., Avadhani, A., Burns, T.C., El-Deiry, W., Wong, D.T., and Rustgi, A.K. (2001). p63 expression is associated with p53 loss in oral-esophageal epithelia of p53-deficient mice. Cancer Res 61, 6467-6473. Thomas, P.E., Wharram, B.L., Goyal, M., Wiggins, J.E., Holzman, L.B., and Wiggins, R.C. (1994). GLEPP1, a renal glomerular epithelial cell (podocyte) membrane protein-tyrosine phosphatase. Identification, molecular cloning, and characterization in rabbit. J Biol Chem 269, 19953-19962. Tureci, O., et al. (1998). Human carbonic anhydrase XII: cDNA cloning, expression, and chromosomal localization of a carbonic anhydrase gene that is overexpressed in some renal cell cancers. Proc Natl Acad Sci U S A 95, 7608-7613. Tusher, V.G., Tibshirani, R., and Chu, G. (2001). Significance analysis of microarrays applied to the ionizing radiation response. Proc Natl Acad Sci U S A 98, 5116-5121. Wang, S.N., Lapage, J., and Hirschberg, R. (2001). Loss of tubular bone morphogenetic protein-7 in diabetic nephropathy. J Am Soc Nephrol 12, 2392-2399. Wilson, C.L., Ouellette, A.J., Satchell, D.P., Ayabe, T., Lopez-Boado, Y.S., Stratman, J.L., Hultgren, S.J., Matrisian, L.M., and Parks, W.C. (1999). Regulation of intestinal alpha-defensin activation by the metalloproteinase matrilysin in innate host defense. Science 286, 113-117. Yano, N., Endoh, M., Fadden, K., Yamashita, H., Kane, A., Sakai, H., and Rifai, A. (2000). Comprehensive gene expression profile of the adult human renal cortex: analysis by cDNA array hybridization. Kidney Int 57, 1452-1459. Yu, J.X., Chao, L., and Chao, J. (1995). Molecular cloning, tissue-specific expression, and cellular localization of human prostasin mRNA. J Biol Chem 270, 13483-13489. Zhang, H., Wada, J., Hida, K., Tsuchiyama, Y., Hiragushi, K., Shikata, K., Wang, H.,Lin, S., Kanwar, Y.S., and Makino, H. (2001). Collectrin, a collecting duct-specific transmembrane glycoprotein, is a novel homolog of ACE2 and is developmentally regulated in embryonic kidneys. J Biol Chem 276, 17132-17139. |
Почка является мультифункциональным органом, играющим критическую роль в селективной элиминации растворимых шлаков, в гомеостазе кислот, электролитов и гормонов, а также в регуляции кровяного давления и эритропоэза. Нефрон - функциональная единица почки, высокоорганизованная структура, состоящая из клеток нескольких различающихся специализированных типов. Ультрафильтрат крови формируется в гломерулах, где существенные жидкости селективно реабсорбируются, в то время как шлаки концентрируются и подвергаются экскреции. Разнообразные функции почки поделены в стереотипном смысле между клетками нефрона и ассоциированных структур. Специализация функций вдоль нефрона отражена в макроанатомии долей почки. Кора почки включает в себя гломерулы, юкстагломерулярный аппарат, проксимальные и дистальные извитые канальцы, где формируется плазма ультрафильтрата и реабсорбируется большинство её компонентов, а медулла (мозговое вещество почки) содержит петлю Генле и собирающие протоки, где ультрафильтрат концентрируется.
Характерный тип функциональной специализации клеток по длине нормального нефрона должен быть связан с соответствующей региональной вариацией паттернов генной экспрессии. Изучение профилей генной экспрессии каждого сегмента может способствовать пониманию работы систем, включённых в функционирование каждой части нефрона. Всестороннее, детальное рассмотрение генной экспрессии будет, кроме того, способствовать пониманию как нормальной физиологии почек, так и её нарушений в случае заболеваний последних. Поддерживая исследования генетических программ, направленные на изучение функционирования нефрона, мы проанализировали тип генной экспрессии в изолированных нормальных гломерулах и разных образцах-пробах нормальной почки, характеризующихся собственной макроанатомической локализацией в долях почек. РЕЗУЛЬТАТЫ Экспрессия генов
Анатомически разные отделы почки были подразделены с помощью иерархического кластерного анализа в соответствии с паттерном общей экспрессии для каждого из 1548 генов с широко варьирующей экспрессией (Рис. 1). 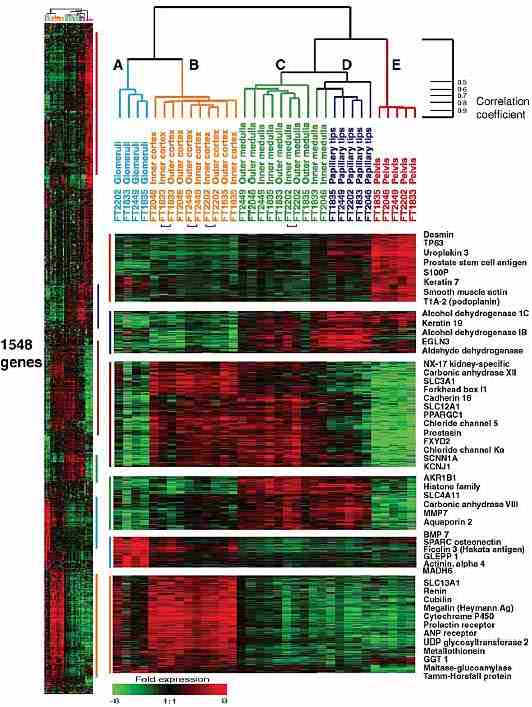 Figure 1: Unsupervised hierarchical cluster. A representation of the variation in expression of 1548 genes in different compartments of five kidneys. Each row represents a single gene. Each column represents a single sample. Green squares indicate transcript levels below the mean; black squares, transcript levels equal to the mean; red squares, transcript levels greater than the mean; gray squares, technically inadequate or missing data. Gene filtering criteria were for four-fold variation from the normalized mean and at least 80% well measured spots. Colored bars adjacent to the clustering table indicate the position of the enlarged images. The color of the bar indicates the type of sample in which the cluster of genes is expressed. Red - pelvis cluster; Dark blue - papillary tip cluster; Brown - tubular cluster; Green - medullary cluster; Turquoise - glomerular cluster; and Orange - cortex cluster. Selected gene names are shown. The full dataset can be explored on the accompanying website: https://www.med.stanford.edu/jhiggins/Normal_Kidney/index.shtml. Главная ветвь дендрограммы отделила пробы из кортекса (включая пробы очищенных гломерул) от образцов медуллы и почечной лоханки. Внутри каждой из этих ветвей можно увидеть последующее разделение: образцы гломерул формируют кластер (ветвь A), который отличается от кластера кортикальных проб (ветвь B). Также пробы лоханки (ветвь E), медуллы (ветвь C) и папиллярных концов (ветвь D) образовывали собственные кластеры. Причём большинство медуллярных проб кластеризовалось в отдельную ветвь, только 2-й и 10-й образцы составляли общий кластер с пробами, полученными их папиллярных концов. Различия между образцами внутреннего и наружного слоёв как для кортекса, так и для медуллы были незначительными. Кортикальный кластер
Более 250 генов экспрессировались преимущественно в коре почки и отличали и эти образцы от других анатомических сегментов. Вероятно, этот характерный паттерн экспрессии генов имел место во многом, благодаря характерным молекулярным свойствам клеток извитых канальцев и гломерул. Специфически экспрессировавшиеся гены гломерул в дальнейшем выявили большее значение в пробах из очищенных гломерул (см. ниже). Проксимальные извитые канальцы являются преимущественным клеточным типом кортекса почки, и не присутствуют ни в каких других её анатомических сегментах. Семнадцать транскриптов, преимущественно экспрессировавшихся в кортексе, кодировали членов семейства растворимых переносчиков (soluble carrier family), включая хлорид натрия, глюкозу, органические ионы и аминокислотные переносчики (amino acid transporters). В этом кластере была обнаружена иодотиронин-деиодиназа I и II типа, что позволило предположить участие кортекса и клеток проксимальных изогнутых канальцев в почечном метаболизме гормона щитовидной железы. Многие гены из кластера кортекса также экспрессировались в тонком кишечнике. Транкрипты некоторые из них участвуют в транспорте питательных веществ, например, SLC2A5 (облегчает перенос глюкозы/фруктозы) и кубилин (встроенный фактор кобаламинового рецептора). Другие, по-видимому, играют роль во внутренней мукозной защите, в том числе фукозилтранфераза 6, которая включена в синтез е-селестин лиганда (lewis антиген группы крови) и может облегчать хоминг (homing) лейкоцитов (Koszdin & Bowen, 1992). Есть группа, участвующая в пищеварении: мальтаза-глюкоамилаза (Nichols et al., 2003), альдолаза B (Cox et al., 1982) и кетогексокиназа (Bonthron et al., 1994). Экспрессия последней группы генов в почка является весьма неожиданной и позволяет предполагать, что данные гены могут обслуживать функцию более высокого порядка, присущую абсорбирующему эпителию.
Гены, обнаруженные в кортикальных пробах, иллюстрируют роль почки в детоксикации. Некоторые из них являются характерными для печени, например, ген аланинглиоксилат-аминотрансферразы. Мутации в этом гене являются причиной гипероксалурии I-го типа, которая обычно приводит к почечной недостаточности (Danpure & Jennings, 1986). Глютатион S-трансфераза A2 является ферментом печёночной ткани, участвующим в детоксикации терапевтических лекарств, канцерогенов, токсинов внешней среды и продуктов окислительного стресса (Mannervik, 1985). Несколько членов семейства UDP-гликозилтрансферазы 2 экспрессировались в этом кластере, продукты этих генов участвуют в элиминации ксенобиотиков и эндогенных токсинов (Radominska-Pandya et al., 2001). Наконец, металлотионовые (metallothionein) IG включены в гомеостаз тяжёлых металлов и предупреждают токсикацию ими (Nath et al., 1988). Экспрессия этой широкой панели генов, включённых в детоксикацию, подчёркивает комплексную роль почек в поддержании клиренса и секреции токсических смесей. Гломерулярный кластер
Для приготовления цельных гломерул со степенью чистоты 95% (Рис. 2) мы использовали метод просеивания (sieving technigue - Misra, 1972).  Figure 2: Isolation of glomeruli from fresh renal cortex shows a high degree of purification. Figure 2: Isolation of glomeruli from fresh renal cortex shows a high degree of purification.
Образцы гломерул дали высоко специфический паттерн экспрессии генов, часть признаков которого также присутствовала в кортикальных пробах. 139 из 1548 генов преимущественно экспрессировались в гломерулах. Среди них были и гены, чья экспрессия уже была установлена в подоцитах. Это гены, кодирующие синтез остеонектина (Floege et al., 1992), актинина-альфа 4 (Kaplan et al., 2000), GLEPPI (Thomas et al., 1994), тесно связывающего белка ZO-1 (Kurihara et al., 1992; Schnabel et al., 1990). Кроме них в гломерулах преимущественно экспрессировались гены гломерулярного эндотелия: TEK-тирозинкиназы (Dumont et al., 1994) и EDGI (Liu et al., 2000), а также мезангиальных клеток - эндоглин (Rodriguez-Barbero et al., 2001; Roy-Chaudhury et al., 1997). Однако для многих транскриптов этого кластера, включая несколько названных выше, прежде экспрессия в гломерулах не отмечалась. В частности для костного морфогенетического протеина 7 (BMP7) описано присутствие в почечных канальцах (Wang et al., 2001), но наши результаты показывают, что в гломерулах его экспрессия выражена даже сильнее. MADH6, также входящий в этот кластер, известен как селективный ингибитор сигнала BMP7, конкурирующий с smad4 за рецептор-активированный smad1 (Hata et al., 1998). Фиколин 3 (hakata антиген) был прежде обнаружен в ткани лёгких и печени (Akaiwa et al., 1999; Fukutomi et al., 1996). Он является геном-мишенью для аутоантител у некоторых пациентов с системной красной волчанкой (Inaba et al., 1990), по-видимому, играя роль в нарушении их гломерулярного аппарата. Не названные и не охарактеризованные гены этого кластера, такие как EST Hs.135335, только идентифицированный в библиотеке кДНК нефробластомы (http://source.stanford.edu), единично экспрессировались в подоцитах, мезангиальных клетках или гломерулярном эндотелии. Кластер почечных канальцев
Мы идентифицировали около 130 генов, для которых была характерна экспрессия в кортексе почки и медулле, но не в изолированных гломерулах или почечной лоханке. Мы предположили, что эти транскрипты относятся к формированию и функционированию эпителия почечного канальца, так как последний является характерной структурой и для кортекса, и для медуллы, но отсутствует в гломерулах и почечной лоханке. Некоторые транскрипты данной группы были уже известны, как экспрессирующиеся в почках. Это простатин (Yu et al., 1995), специфичный для почек мембранный белок NX-17 (Zhang et al., 2001), CLCNKA (Matsumura et al., 1999), FXYD2 (Arystarkhova et al., 1999), SLC12A1 (Simon et al., 1996), SCNN1A (Canessa et al., 1994) карбоангидраза XII (Tureci et al., 1998). Последние четыре гена являются мишенями для широко используемых диуретиков и демонстрируют потенциальную возможность их использования в будущем получении фармакологических агентов прямого действия.
Некоторые из не охарактеризованных генов данного кластера представляют большой интерес. Например, ESTs Hs.144472, Hs.155290, Hs.135787, Hs.126246, Hs.155747, Hs.128408 представлены клонами, которые были изолированы исключительно из библиотек кДНК почки (http://source.stanford.edu). По-видимому, они являются специфическими генами почки с пока не выявленными функциями. Медуллярный кластер
В некоторых пробах мозгового слоя почки и папиллярных концов наблюдалось перекрывание, с концами вместе кластеризовались и две пробы внутренней части медуллы. В частности, папиллярные концы формировали один кластер транскриптов совместно с медуллой и второй - с почечной лоханкой (Figure 1). Данное перекрывание является свидетельством того факта, что клеточные компоненты папиллярных концов имеют свойства клеток медуллы и почечной лоханки. Или же часть компонентов папиллярных концов (интерстиций и канальцы) также наличествует в мозговом слое почки (такие, как уротелий на внешней поверхности сосочка), тогда как другие присутствуют в клетках почечной лоханки. Последняя возможность предполагается в связи с наблюдением экспрессии нескольких кератинов и в ткани почечной лоханки, и в папиллярных концах. Эти кератины постоянно экспрессировались в эпителии. С другой стороны, аквапорин 2 (ген, экспрессирующийся в собирающем протоке) обнаруживается в медуллярных образцах, что служит указанием на возможно непрерывный, последовательный паттерн экспрессии вдоль нефрона. Сходное перекрывание экспрессии установлено для генов кортекса и медуллы. Многие из них, возможно, представляют широко экспрессирующиеся гены почечных канальцев (такие, как ген Na-K-АТФазы), так как канальцы являются компонентом, присутствующим и в кортексе, и в медулле. Но в гломерулах и ткани почечной лоханки канальцы отсутствуют.
Группа из 58 генов более высоко экспрессировалась в мозговом слое почки, чем в папиллярных концах. Она включала несколько ожидаемых генов, например таких, как аквопорин 2. Транскрипты двух генов в медуллярном кластере могли играть роль в защите слизистой оболочки: Fc-фрагмент белка, связанного с IgG, и металл-протеиназа 7 матрикса, которая вовлечена в процесс излечивания ран (Lu et al., 1999), а также может регулировать активность защиты слизистой оболочки кишечника (Wilson et al., 1999). Предполагается, что механизмы, работающие в последнем случае, могут представлять общую стратегию защиты от микробов в эпителии разных органов. Кластер папиллярных концов
Транскрипты 102 генов папиллярных концов дали профили, частично совпадающие с таковыми для медуллы и почечной лоханки, что, вероятно, отражает совпадение клеточных типов этих анатомических районов. Некоторые из генов данного кластера имеют непосредственное отношение к unanticipated функциям в папиллярных концах. Ген SLC21A3 опосредует натрий-зависимый клеточный перенос органических ионов в печени, включая жёлчные кислоты, бромсульфофталеин и некоторые стероидные компаунды (Kullak-Ublick et al., 1995). C1 - член семейства 1 альдо-кеторедуктазы также входит в этот кластер, по-видимому, участвуя в транспорте жёлчных кислот (Stolz et al., 1993). Несколько генов, включённых в метаболизм алкоголя (таких, как алкоголь-дегидрогеназа IB и IC, член A3 семейства альдегид-дегидрогеназы 1), также высоко экспрессировались в папиллярных концах. В то время как экспрессия алкоголь-дегидрогеназы I была прежде установлена в почечных канальцах человека (Buhler et al., 1983), наши результаты по нескольким генам, включённым в метаболизм алкоголя, отводят важную роль в данном процессе папиллярным концам. Кроме того, локализация этих генов в медулле позволяет предположить, что в условиях гипоксии мозгового слоя почки продукты метаболизма алкоголя могут продуцироваться в увеличенном количестве. Кластер почечной лоханки
Довольно большой кластер генов экспрессировался преимущественно в ткани почечной лоханки, свидетельствуя о различиях между этой тканью и другими частями почки. Многие транскрипты этого кластера, такие как гамма-2-актин, кавеолин 1, кальпонин 1, тропомиозин 2, киназа лёгкой цепи миозина и актин-связывающий белок 280, служат отражением гладкой мускулатуры почечной лоханки. Другие гены этого кластера, включая уроплакин 1В и 3 (Lobban et al., 1998), а также антиген стволовых клеток простаты (Amara et al., 2001), экспрессируются уротелием почечной лоханки. Продукты третьей группы генов участвует в защите уротелия от неопластической трансформации. В неё входят глутатион-пероксидаза 2, защищающая желудочно-кишечный тракт от органических гидроперекисей (Chu et al., 1993), и TP63, который может участвовать в индукции апоптоза и ингибировании роста в p53-дефицитных клетках (Suliman et al., 2001). Внутренние слои кортекса и медуллы в сравнении с их внешними слоями
Паттерны экспрессии генов внешнего и внутреннего слоёв кортекса показали высокую степень сходства, не разделяемую методом иерархического кластерного анализа. У одних и у тех же пациентов три из пяти проб внешнего слоя кортекса дали генную экспрессию, наиболее сходную с таковой для внутреннего слоя. 49 генов (Webtable I) экспрессировались во внутреннем слое кортекса сильнее, чем во внешнем (false significant number: 4). Ни один ген не экспрессировался во внешнем слое кортекса сильнее, чем во внутреннем. Десять генов (Webtable II) экспрессировались во внутреннем слое медуллы сильнее, чем во внешнем (false significant number: 3). Будущие исследования помогут установить функциональное значение этих генов. Иммуногистохимия
Для иммуногистохимического исследования были выделены 5 транскриптов родственных белков нефрона. Установлено, что транскрипционный уровень SERP1 (secreted frizzled-related protein 1) выше в коре почки, чем в её мозговом слое. Сходный паттерн экспрессии ясно виден (на левой панели Figure 3) у белка в кортексе и медулле. При высоком разрешении (правая панель) видно, что белок SERP1 экспрессируется в проксимальных извитых канальцах гораздо сильнее, чем в дистальных. Сходным образом, транскрипционный уровень SLC9A3R1 (solute carrier family 9, isoform 3 regulatory factor 1) был наибольшим эпителии проксимальных канальцев (Webfigure 2). Этот белок первоначально характеризовался как присущий коре головного мозга, а затем был установлен высокий уровень его экспрессии в почке (Reczek et al., 1997). По-видимому, он важен в поддержании полярности клетки (Bonilha & Rodriguez-Boulan, 2001).
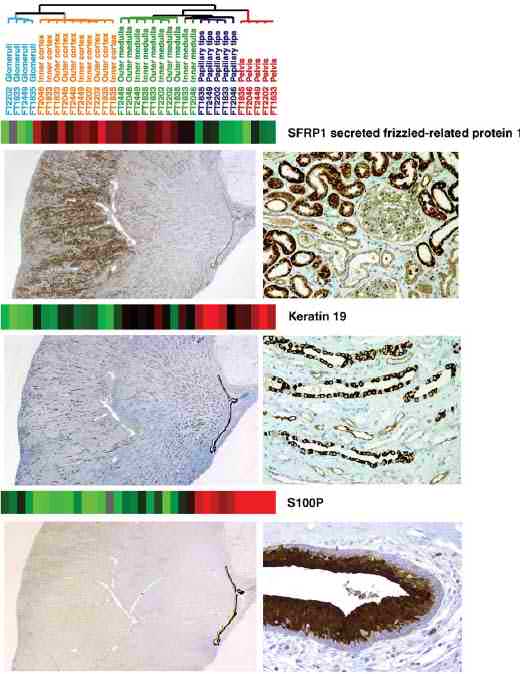 Figure 3: Correlation of mRNA with protein expression by immunohistochemistry. Messenger RNA expression levels for three genes, SFRP1, keratin 19, and S100P have been taken from figure 1 together with the figure 1 sample dendrogram. Immunohistochemical stains for each of these genes are shown below the mRNA data. Иммуногистохимия показала наличие транскрипта кератина 19 в канальцах (Figure 3), но максимальная концентрация его отмечена в папиллярных концах. В клетках висцерального эпителия (подоцитах) была отмечена высокая экспрессия белка SPARC, как сообщали и другие авторы (Floege et al., 1992). В почечной лоханке нами был установлен высокий уровень транскриптов S100P. Иммуногистохимически было подтверждено, что этот белок присущ только эпителию почечной лоханки и совершенно не встречается в тубулярном эпителии. Данные об экспрессии S100P в уротелии были получены впервые в нашей работе. Обсуждение
Результаты нашего исследования показывают, что различающиеся макроскопически компартменты почечных долей являются отражением различий и репродуктивных типов генной экспрессии. Каждый из компартментов давал высоко специфичный тип экспрессии генов. В то время как пара образцов от одного и того же пациента была наиболее сходной в плане наблюдавшейся экспрессии, вариация последней в каждом компартменте образцов от разных пациентов была гораздо меньше, чем та, что наблюдалась для опухолей почки (Higgins et al., 2003). Эта согласованность характеристик профилей генной экспрессии в каждом районе или субструктуре почки продемонстрирована плотным группированием сходных образцов в иерархической кластеризации.
При рассмотрении различающихся анатомически субкомпартментов почки мы расширили результаты, прежде полученные другими исследователями для 9-ти образцов кортекса почки взрослого человека (Yano et al., 2000). Некоторые наши результаты совпадают, в частности - высокий уровень экспресии для генов, кодирующих перенос ионов / транспорт белков, и для металлотионовых генов. Однако в нашей работе был установлен более высокий уровень экспрессии рибосомных генов в медуллярных образцах по сравнению с образцами кортекса. Наши результаты добавили детальный профиль экспрессии генов в мозговом слое почки, папиллярных концах и почечной лоханке и позволили провести сравнение экспрессии генов в кортексе с другими компартментами почек. Включение в анализ изолированных гломерул весьма полезно, так как этот кластер может быть связан с генными и иммунными дефектами гломерулярного аппарата, уникальными для почки.
Наша попытка классифицировать экспрессию генов в нормальном нефроне имеет некоторые ограничения. Нефрон имеет извилистое направление в почечной доле, в кортексе локализованы его проксимальные и дистальные извитые канальцы. Кроме того, в состав нормальной почки, помимо эпителия нефрона, входят клетки нескольких типов. Таким образом, гены, экспрессия которых идентифицируется в кортексе, могут экспрессироваться и в эпителии канальцев, в нескольких различающихся типах гломерулярных клеток, в перитубулярных капиллярах или более крупных сосудах, в нормально представленных клетках интерстиция (Lemly & Kriz, 1991) или в лейкоцитарном инфильтрате и дендритных клетках.
Несмотря на эти ограничения, возможности использованной технологии позволили установить гены, имеющие значение как для нормальной физиологии почки, так и для развития почечных заболеваний, и разместить их в индивидуальных кластерах. Например, гломерулярный кластер содержит 196 генов и включает в себя такие, как GLEPP1, ZO-1, актин-альфа 4 и остеонектин. Из генов, экспрессирующихся в гломерулах, некоторые задействованы в гломерулонефропатиях. "Переоткрытие" нами этих важных генов показывает, что многие не охарактеризованые гены в этом кластере будут отнесены либо к контролю нормального функционирования гломерул, либо к возникновению в последних патологических процессов. Один новый кандидатный ген - фиколин 3 (Hakata антиген), сывороточную реакцию на который дают пациенты с красной волчанкой. Экспрессия его до сих пор была установлена в печени и лёгких. Возможно, наиболее интересными открытиями наших экспериментов являются панели экспрессии явно родственных, но неожиданных генов. Экспрессия в коре почки генов детоксикации, в папиллярных концах - генов метаболизма желчи, в мозговом слое почки - генов защиты слизистой кишечника позволяет предположить, что почка может играть роль в биологических процессах, которые прежде считались исключительной прерогативой других органов. Напротив, может быть, что в почке эти гены являются основой функций, отличающихся от тех, которые прежде были описаны для них в других органах.
|