Axonal neuregulin-1 regulates myelin sheath thickness
Science. 2004 Apr 30; 304 (5671). P.700-703. | |
|
|
   Thickened myelin sheath due to multiple layers of folded myelin 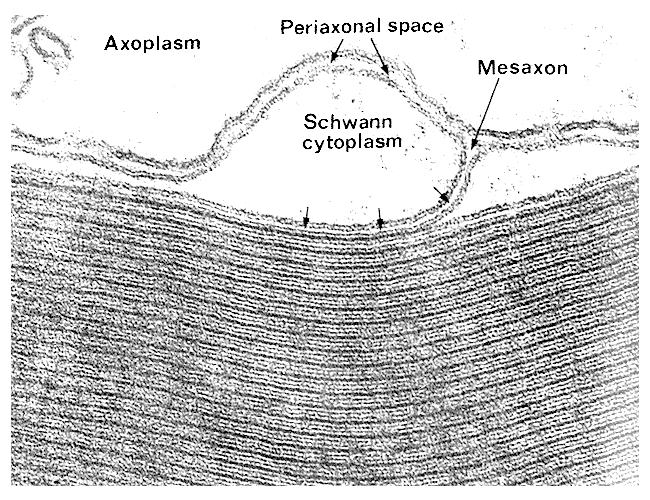
Имеется определенная и прогнозируемая связь между диаметром аксона и толщиной окружающего его миелина. Авторы представили доказательства того, что neuregulin-1 (Nrg1),продуцируемый аксонами, передает информацию о толщине миелиновой оболочки Шванновским клеткам (Schwann cells).
Скорость, с которой потенциал действия распространяется по аксону, в основном зависит от толщины миелиновой оболочки и от диаметра аксона – двух взаимосвязанных факторов. Контроль за скоростью проводимости крайне важен для функционирования нервной системы. Предполагается, что толщина миелина может контролироваться взаимодействиями между лигандами, продуцируемыми аксонами, и рецепторами в миелинизированной глиии. Авторы предположили, что таким аксонным лигандом может быть Nrg1, взаимодействующий с ErbB рецепторами на Шванновских клетках.
Ген Nrg1 экспрессируется в нейронах седалищного нерва мышей, а ErbB2 и ErbB3 экспрессируются Шванновскими клетками. Если взаимодействия между Nrg1 и ErbB рецепторами контролируют толщину миелина, то изменения в количествах лиганда или рецептора могут менять толщину миелиновой оболочки. Когда были получены компаундные гетерозиготные мыши со сниженной дозой гена Nrg1 и ErbB2, то оказалось, что миелин в седалищных нервах мышей был значительно тоньше, чем у контрольных животных. Хотя внешне мыши выглядели нормально, скорость проводимости в их нервах была значительно снижена, даже если размеры аксонов не изменялись.
Для выяснения причин снижения миелинизации, авторы исследовали мышей со сниженной дозой гена Nrg1 или ErbB2. У мышей гетерозиготных по ErbB2 количество миелина оказалось нормальным, однако у Nrg1 гетерозигот миелин был такой же толщины, что и у компаундных гетерозигот. Вероятно, экспрессия Nrg1 контролирует толщину миелина.
Для дальнейшей проверки данной гипотезы авторы сконструировали мышей, у которых Nrg1 сверхэкспрессировался под контролем мышиного промоутера Thy1.2 так, что избыток Nrg1 специфически экспрессировался в постнатальных моторных нейронах и нейронах дорсальных корешковых ганглиев. У этих мышей по сравнению с мышами дикого типа периферические нервы были гипермиелинизированы.
Известны три изоформы Nrg1. Оказалось, что наблюдаемый авторами эффект специфичен для изоформы Nrg1 type III. Мыши, у которых была сверхэкспрессия Nrg1 type III, характеризовались гипермиелинизацией, однако у мышей со сверхэкспрессией Nrg1 type I такого дефекта не наблюдали (хотя у них были некоторые аномалии миелина в центральной нервной системе). Снижение экспрессии Nrg1 type III изоформы вызывало гипомиелинизацию, указывая на то, что Nrg1 type I и II не могут компенсировать отсутствие изоформы Nrg1 type III.
В итоге авторами была предложена модель, согласно которой Nrg1 type III продуцируется в зависимости от диаметра аксона. Количество Nrg1 обусловливает количество сигнала через ErbB рецепторы Шванновских клеток и это контролирует уровень миелинизации каждого аксона. Возникает, однако, вопрос – каким образом образование Nrg1 контролируется количественно и какой сигнальный путь ответственен в результате за предписанную толщину миелина? См. также:  | Electron micrograph and schematic drawing of myelin and its major membrane proteins и |