 | WMZ: Z191701361450 WMR: R209318204033 |
Без рекламы только Браузер Uran (скачать )
| WMZ: Z191701361450 WMR: R209318204033 |
After the Holy Grail: Establishing a Molecular Basis for Mammalian Olfaction Cell V.116, N0 2, P. 329-336, 2004. | ||
 | Сайт по обонянию
Восприятие запах людьми очень сильно отличается между индивидами, описаны огромные вариации в восприятии как по интенсивности, так и приятности (pleasantness) данного запаха. Напр., androstenone (5a-androst-16-en-3-one), запах стероида, происходящего из тестостерона, по разному воспринимается разными индивидами как противный ("пота, мочи"), приятный ("сладкий, цветочный") или не имеющий запах1-3. Сходная изменчивость в восприятии запаха наблюдается и для некоторых др. запахов4-6. Механистические основы изменчивости в воприятии запахов между индивидами неизвестн. Исследовали, может ли генетичекая изменчивость у людей в генах рецепторов одорантов объяснить частично изменчивость в восприятии между индивидами7,8. Было установлено, что рецептор запаха у человека, OR7D4, избирательно активируется in vitro с помощью androstenone и родственного запаха стероида androstadienone (androsta-4,16-dien-3-one) и не реагирет на панель из 64 др. запаха и 2 растворителя. Наиболее распространенный вариант этого рецептора (OR7D4 WM) содержит два не-синонимных single nucleotide polymorphisms (SNPs),возникших в результате двух аминокислотных замен (R88W, T133M; отсюда 'RT'), которые серьезно нарушают функцию in vitro. Индивиды с RT/WM или WM/WM генотипами, как группа менее чувствительная к androstenone и androstadienone, находит оба запаха не такими неприятными, как RT/RT группа. Генотипическая изменчивость в OR7D4 объясняет существенную пропорцию значимости (приятности или неприятности) и интенсивности вариабельности в восприятии этих стероидных запахов. Полученные результаты демонстрируют впервые связь между функцией рецепторов запахов у людей in vitro и восприятием запаха.  Рисунки к статье | | The Identification of Odorant Receptors 10 миллионов сенсорных нейронов формируют сенсорный эпителий носа, который обеспечивает инициальное восприятие и передачу пахучих стимулов. Дифференциальная реакция индивидуальных клеток на единственный запах из спектра от 10 до тысячи тысяч химических структур, которые различает человек является основой чувствительности и специфичности этой системы. Молекулярно генетические различия этих нейронов, большое количество типов обонятельных рецепторов и профили реакции каждого рецепторного белка и предопределяют физиологические свойства таких клеток. Ранние анатомические исследования показали, что инициальные сигналы, генерируемые с помощью обонятельных рецепторов нейронов эпителия, подвергаются экстенсивной переработке на инициальной релейной станции, в обонятельных луковицах (olfactory bulb (OB)). Несколько тысяч отдельных гломерул воспринимают аксональные сигналы из эпителия, а также с дендритных и аксональных проекций от множественных типов клеток внутри луковиц. Эти гломерулы являются функциональными единицами процессинга обонятельной информации.
Рецепторы, ответственные за инициальную трансдукцию стимулов запаха составляют ключевой элемент развивающейся сети. Анализ этих рецепторов фундаментально изменил наше отношение к характеристике обонятельной системы. Идентификация генов, кодирующих обонятельные рецепторы, стала возможной благодаря анализу сигнальных систем. В частности, характеристики компонентов зрительной системы млекопитающих, с её горсткой рецепторов, инициирующих с G-протеином связанный сигнальный путь, представляющим парадигм для кодирования информации по интенсивности и качеству сенсорных стимулов. Более того, структурное сходство свет-воспринимающих опсинов и множественных рецепторов нейротрансмиттеров и гормонов расширили небольшие области законсервированных аминокислотных последовательностей позволили идентифицировать новые семейства G protein coupled receptors (GPCRs)
Действительно, было выявлено участие G белками опосредованных путей в обонянии млекопитающих и насекомых и была подтверждена специфическая роль цАМФ и IP3 , соотв., в двух филетических типах. Присутствие специфических для обоняния трансдукционных белков, которые обеспечивают нижестоящие сигнальные события, строго указывает на то, что передача сигналов цАМФ является общим путём для трансдукции запахов, но мало что даёт для идентификации рецепторов, сложности семейства рецепторов и паттерна их экспрессии в сенсорном эпителии.
Исходная публикация (Buck and Axel, 1991), в которой были идентифицированы гены, кодирующие odorant receptors (ORs), базировалась на гипотезе, что большое семейство генов, достаточное для детекции тысяч химических структур, д. в результате давать амплифицированные фрагменты ДНК, представленные множественными молекулярными видами. Последующая характеристика индивидуальных клонированных изолятов выявила новое семейство связанных кДНК, которые обладали общими признаками с др. известными GPCRS. Свойства этого семейства генов удовлетворяли и некоторым др. критериям, предсказанным для обонятельных рецепторов. А именно, д.б. сотни родственных рецепторов, кодируемых в геноме млекопитающих, а экспрессия этих рецепторов д.б. ограничена обонятельным эпителием. Genomic Organization and Evolution of ORs Выявление и дискриминация 10 тысяч или более структурно отличных соединений представляет собой фундаментально более значительную задачу, чем вопрос опсинов, захватывающих фотоны, и отличающиеся только по энергии последних. Структура GPCR рецепторов представляет собой ужасное препятсвие для иммуноглобулин-подобного механизма генерации гипервариабельных областей и чрезвычайного разнообразия рецепторных белков. В GPCRs аминокислоты, которые вносят вклад в связывание лиганда, распределены по нескольким трансмембранным доменам, и множественные события рекомбинации, каждое из которых склонно к генерации дефектного белка, необходимы для создания разнообразия в этой системе. Итак, экспрессия разнообразных, но из вполне определенного семейства, индивидуально кодируемых рецепторов даёт в результате большие нейрональные популяции, экспрессирующие идентичные рецепторные белки. Последующая организация connectivity и circuitry обонятельных луковиц для процессинга обонятельной информации, по-видимому, существенно зависит от этого факта.
OR гены необычно компактны, весь белок кодируется единственным экзоном. Это семейство генов составляет от 1% до 3% генов от всего количества генов, выявленных у млекопитающих. Наблюдаются драматические различия в частотах псевдогенов у грызунов и человека, с небольшим процентом дефектных генов у мышей и до 2/3 у людей, т.е. генов, лишенных открытой рамки считывания. Сравнение с др. геномами подтверждает, сто мы потеряли большинство из наших рецепторных генов, когда стали двуногими и наш нос удалился от земли. Потеряны целые подсемейства во время дивергенции этих видов, тогда как др. были чрезвычайно увеличены.
Ряд сил управляет динамикой эволюции семейства генов OR. Организация генов OR, распределение практически по всем хромосомам вносят вклад в быструю эволюцию. Высоко гомологичные рецепторы образуют тесные кластеры в геноме и часто располагаются в субтеломерных областях, где генетические изменения наиболее часты. Кроме того, неравная рекомбинация между родственными рецепторами в нутри геномного кластера м. вносить вклад в экспансию специфических подсемейств.
Идентификация и характеристика ORs млекопитающих ведет к выявлению аминокислотных последовательностей, которые обнаруживаются в ORs и др. позвоночных. Интересно, что водные виды обладают характерными белковыми мотивами, которые общи субнабору рецепторов у Xenopus, которые экспрессируются во время водной фазы их развития. Эти мотивы обнаруживаются также в субнаборах ORs млекопитающих, хотя лиганды для этих рецепторов неизвестны. После метаморфоза Xenopus экспрессируется второй класс рецепторов, более сходный с теми, что имеются у дышащих воздухом животных
Сходство последовательностей ORs не распространяется на беспозвоночных. Основные вторичные мессенджеры, используемые этими организмами, отличаются, а в некоторых случаях ограниченное число рецепторных клеток у простейших организмов обладает фундаментально иной молекулярной логикой ощущения запахов. Выясненеие молекулярных основ обоняния Drosophila шло параллельно с таковым у млекопитающих. Анализ хемосенсорных мутантов у C. elegans привел к идентификации семейства GPCRs, которое определяет летучие аттрактанты и репеленты. Эти исследования выявили относительно большие количества генов ORs несмотря на вариации в размерах геномов. Это указывает на то, что величина репертуара ORs у всех организмов представлена примерно 100 - 1000 генами, способными воспринять весь широкий сегмент запахов пространства. Related Expression of Individual ORs C помощью гибридизации in situ индивидуальных OR генов с обонятельным эпителием было установлено, что каждый рецептор экспрессируется только в небольшой фракции обонятельных нейронов. Было предположено, что каждая клетка экспрессирует только один или немного рецепторных генов. С помощью PCR методов было подтверждено, что каждая клетка экспрессирует только один рецептор, хотя недавние эксперименты усовершенствовали это One OR - One Neuron правило.
Ранние исследования показали, что OR, экспрессируемый в каждой клетке, происходит только с одного из двух имеющихся аллелей в геноме, а выбор, какой из аллелей будет задейстован случаен. Следовательно, популяция, экспрессирующая этот ген, будет подразделена на две субпопуляции по экспрессируемому аллелю. Моноаллельная экспрессия OR генов коррелирует с асинхронной репликацией OR локусов во всех клетках мышей. Согласно одной модели два аллеля одного рецептора выбираются для экспрессии и один из них молчит с помощью 'allelic exclusion'б то согласно современным моделям каждый из 2000 OR аллелей, имеющийся в геноме мыши функционирует независимо без специфического взаимодействия двух аллелей данного рецепторного локуса. Выбор аллеля, является механизмом, гарантирующим, что только один OR будет экспрессироваться в каждой клетке. Чтобы понять феномен асинхронной репликации, общий всем OR локусам, и его роль в моноаллельной экспрессии, нужны генетические системы с нарушениями этого процесса.
Нейроны, экспрессирующие индивидуальные OR гены, не распределяются униформно в сенсорном эпителии. Каждый рецептор компартментализован в одной из 4-х зон экспрессии, организованных вдоль дорсо-вентральной оси. Выясняется довольно сложный паттерн оккупации субсемействами рецепторов характерных доменов, которые частично или полностью перекрываются с др. рецепторами. Рецепторы одного и того же подсемейства перекрестно гибридизируются в этих in situ экспериментах и таким образом предопределяется их зона экспрессии, но эта перекрёстная реактивность м. затемнять тонкую структуру паттерна экспрессии рецептора.
Механизмы, которые контролируют экспрессию OR, изучали с помощью геномной и генетической техники. Хотя OR гены млекопитающих обнаруживают заметную консервацию организации с одним или немногими 5энетранслируемыми экзонами, сопровождаемыми интроном в 3-10 п.н. и единственной кодирующей областью экзона. Гомология последовательностей ДНК в промоторах чрезвычайно ограничена во всех, за исключением очень близких рецепторов. В двух исследованиях ДНК последовательности в несколько т.п.н. выше места старта транскрипции оказались достаточными для управления стохастическим, зона-ограниченным паттерном экспрессии, характерными для генов рецепторов. Промоторные области, самое малое в 500 п.н., поддерживают большинство аспектов регулируемой экспрессии.
Имеющиеся сайты связывания для О/Е белков, указывающих на транскрипционную регуляцию др. специфичных для обоняния генов, являются единственными законсервированными элементами, идентифицированными в области, достаточной для экспрессии репортерного гена. Интересно, что эти О/Е сайты связывания м.б. идентифицированы во многих промоторах OR, а значит они м. представлять собой общий цис-элемент, существенный для экспрессии рецептора. В одном рецепторном локусе последовательность ДНК в 80 т.п.н. выше гена OR оказалась необходимой для экспрессии всех рецепторов геномного кластера. Эта регуляторная последовательность законсервирована у людей и мышей. Являются ли эти удалённые регуляторные элементы уникальными для этого кластера рецепторов или напротив являются общей характеристикой многих рецепторных генов, м. установить только при анализе дополнительных OR локусов. The Role of OR Protein in Regulation of Receptor Expression Недавно трансгенные модели выявили механизм обратной связи, который гарантирует экспрессию одного OR в каждом нейроне. Экспрессия одиночного функционального OR в каждом из нейронов доказана. Однако, когда OR промотор управляет экспрессией репортера, который замещает OR кодирующей области, то дополнительные ORs коэкспрессировались в таких клетках. Аналогичные конструкции, в которых OR последовательность была представлена в трансгенной мРНК, но которая не транслоцировалась в белок, подтверждают, что это OR белок обеспечивает сигнал обратной связи, чтобы предупредить экспрессию дополнительных OR генов. Важным следствием этих исследований явилось то, что активация OR псевдогена не способна инициировать подобную петлю обратной связи и делает возможной экспрессию второго функционального OR белка в этих клетках.
Идентификация ОR белка, как критического компонента, регулирующего экспрессию OR, представляет новые пути для исследований. Компоненты пути, которые передают сигналы от OR белка на плазматическую мембрану к OR локусам в ядре, неизвестны.
Пути, которые используются для распространения сигнала обратной связи от OR белка, м. также объяснить характерную ламинарную организацию, наблюдаемую для экспрессии некоторых рецепторов. Самые ранние предшественники обонятельных нейронов располагаются вблизи базальной ламины эпителия. Эти клетки мигрируют апикально, как только они дифференцируются и начинают экспрессировать ORs. Если активация индивидуального рецепторного локуса регулируется во времени при такой миграции, то тот же самый сигнал, который ограничивает экспрессию дополнительных рецепторов, м. также вызывать прекращение OR-экспрессирующей клеткой апикальной транслокации и дифференцировку внутри ограниченной слоя (lamina). Ligand Recognition and Specificity Прямая демонстрация, что один член семейства м. инициировать физиологическую реакцию на одорант, впервые была осуществлена путём повторного внесения клонированного рецептора ORI7 в эпителий. Способность octanal и heptanal, но не альдегидов с более короткой цепью, активировать этот рецептор впервые позволила измерить избирательность ORs. Последующие структурно-функциональные исследования выявили спектр одорантов, которые м. активировать этот рецептор.
Альтернативный подход к идентификации рецепторов, способных отвечать на определенные одоранты, связан с молекулярным клонированием ORs, экспрессирующихся на диссоциированных обонятельных нейронах в ответ на определенные пахучие стимулы. В вариантах таких подходов генерация бицистронных, OR-репортёр-нагруженных клеток, позволила сфокусироваться на идентификации лигандов. которые м. активировать меченные клетки. Такой анализ создаёт более пригодную и воспроизводимую систему для идентификации лигандов для идентифицированных рецепторов. Использование методов intrinsic optical imaging позволило визуализировать активные области обонятельных луковиц (Bolluscio , Katz, 2001; Meister, Bonhoeffer, 2001), это могло бы создать карту, пригодную для молекулярной характеристики рецепторов, экспрессируемых конвергирующими первичными нейронами.
Ограниченные количества рецепторов, функционально экспрессируемых в гетерологических клеточных линиях, и исследования их экспрессии указывают на то, что слияние ORs с N концом дивергентных GPCR способствует собственно экспрессии и локализации в плазматической мембране. Неужели эта неспособность собственно транслоцировать OR белок на клеточную поверхность в гетерологических системах является результатом отсутствия существенных кофакторов, присутствующих только в обонятельных нейронах, или, напротив, существуют пути в гетерологических клетках, которые секвестрируют рецепторный белок во внутриклеточном компартменте.
Существование специфических anosmias, неспособности ощущать определенные одоранты, у людей и мышей, как полагали, возникают благодаря функциональному разнообразию ORs внутри соотв. репертуара рецепторов (Amoore, 1967; Wysocki et al., 1977). У людей и мышей специфические anosmias обнаруживают строгую генетическую зависимость (Wysocki, Beauchamp, 1984). Одна специфическая anosmia, редуцирующая способность воспринимать запах isovaleric кислоты (IVA) некоторыми линиями мышей, наследуется как простой Менделевский признак, картируемый в небольшой области хромосомы 4 (Griff, Reed, 1995). Кластер мышиных ORs, картируемых внутри области в 0.5 сМ, которая содержит и локус IVA1, который является лучшим кандидатом для кодирования видо-специфической изменчивости в чувствительности к IVA (Zhang, Firestein, 2002).
Отсутствие четко доказанных взаимодействий одорант-рецептор не позволяет решить вопрос о распознавании одорантов и трансдукции. Рецепторы д.б. настроены широко, способны связывать широкий спектр химических структур, а идентификация лиганда будет производной от разных величин перекрывающихся реакций рецепторов. Возможно также, что существует более узкая настройка рецепторов в результате будет действовать система, в которой идентификация запаха будет определяться с помощью присутствия или отсутствия активности в определенных популяциях рецепторов. Хотя современные исследования свидетельствуют в пользу первого сценария, однако нельзя решить этот важный вопрос в отсутствие генетически идентифицированных вариантов рецепторов, которые бы проявлялись разным поведением или физиологическими реакциями. The Role of ORs in Axonal Pathfinding Способность селективно меченных клеток экспрессировать определенные рецепторы и следовать траектории своих аксонов представляет один из наиболее важных побочных продуктов экспериментов Buck и Axel. Оказалось возможным манипулировать клонированными рецепторами на зародышевой линии мышей, что выявило беспрецендентную точность нахождения пути аксонами и прямую роль OR белка в определении соотв. мишеней.
Обонятельные нейроны, экспрессирующие определенный рецептор обнаруживают удивительную конвергенцию своих аксонов на паре медиальных и латеральных гломерул в обонятельной луковице. Эта конвергенция наблюдается как при гибридизации in situ, которая выявляет мРНК OR в окончаниях аксонов, так и при визуализации репортером нагруженных индивидуальных OR аллелей. У генетически изменённых мышей, одиночное бицистронное сообщение кодирует рецептор и модифицированный GFP или lacZ белок, которые локализуются в аксонах и нервных окончаниях. Хотя имеется некоторая вариабельность в точности нахождения, эти исследования представили строгие доказательства, что проекции к дополнительным гломерулам не вносят вклада в схемы (circuitry), которые усовершенствуют и перерабатывают сенсорную информацию в луковицах. Создание специфических нагруженных репортёрами линий мышей является одним из наиболее мощных инструментов для выяснения организации и функции обонятельной системы млекопитающих.
Обонятельная система млекопитающих подвергается полной топографической реорганизации, т.к. первичные нервные волокна проецируются к своим мишеням. Аксоны в объединённых пучках, которые располагаются под базальной ламиной первоначально располагаются в соответствии со своей областью эпителия, из которой они происходят. После контакта с обонятельной луковицей, эти аксоны выходят из пучков и повторно рассортировываются, объединяясь с аксонами, экспрессирующими тот же самый рецептор и конвергируют, чтобы создать гломерулы в стереотипическом положении в луковице. Наблюдение, что аксоны, экспрессирующие идентичные рецепторы, экстенсивно объединяются задолго до того как они достигнут своих мишеней в луковице указывает на то. что само-ассоциация м.б. важным компонентом в организации этой системы. Возникающая в результате топическая карта рецепторов фундаментально отличается от топографической репрезентации первичных сенсорных структур, поддерживаемых с помощью проекций периферических нейронов в ЦНС для большинства др. сенсорных систем.
OR белки непосредственно участвуют в конвергенции в определенные гломерулы обонятельных луковиц. Эксперименты, в которыех кодирующая область одного рецептора замещалась др. в его нормальном генетическом положении оказались достаточными в большинстве условий, чтобы перенаправить эти аксоны в места. ожидаемые для внесённых рецепторов. Аминокислотные последовательности в рецепторах, которые вносят вклад в само-ассоциацию и выбирают мишень д. идентифицироваться с помощью сходных механизмов. Интересно, что аксоны от клеток, экспрессирующих модифицированный аллель, нагруженный репортером, конвергируют в отдельные гломерулы в эктопических позициях, если замещены рецепторами от разных зон. Это м. отражать просто топографическое ограничение вентральных аксонов, достигших своих соотв. дорсальных мишеней или барьеры на пути миграции в комбинации с их очевидной склонностью объединяться в пучки с др. эктопическими аксонами, экспрессирующими тот же самый рецептор.
ORs обеспечивают важные механизмы, которые предопределяют нахождение мишеней в обонятельных луковицах, но дополнительные поверхностные белки обеспечивают притягивающие и отталкивающие сигналы, чтобы создавать необходимые условия для миграции и инннервации. Один класс находящих пути молекул, ephrins и их рецепторов, дифференциально экспрессируются в отдельных популяциях рецепторов и м. предопределять некоторые аспекты формирования передне-заднего паттерна. Избирательная избыточная экспрессия ephrin-A5 в субнаборе нейронов, содержащих один OR ведет к конвергенции этих нейронов в более переднюю область ОВ. Сходным образом, генетическая элиминация молекул ephrin A направляет те же самые аксоны кзади.
Конвергенция аксонов из клеток, содержащих те же самые ORs обеспечивает и сегрегацию этих проекций от аксонов из др. клеток, которые м. экспрессировать рецепторы почти с идентичными аминокислотными последовательностями. Эта почти безупречная конвергенция м.б. нарушена разными факторами. Часто экспрессия репортёров с бицистронных мРНК приводит к тому, что аксоны достигают соседних гломерул, а не тех, которые иннервируются аксонами с нативными рецепторами. Такие аксоны м. б. неспособны интегрироваться с нативными гломерулами из-за изменений в уровне OR белка или снижения нейрональной активности. Модель зависящей от активности конкуренции обонятельных нейронов за мишени в гломерулах и за клеточную выживаемость указывает на то, что проекции аксонов к их конечным мишеням надёжно предопределены (hardwired), но что пахучая среда м. вносить уточнения в соединения и расхождения аксонов, экспрессирующих один и тот же рецептор.
Способность обонятельной системы позвоночных собирать и правильно перерабатывать сенсорную информацию даже удивительна в свете постоянного замещения первичных обонятельных нейронов в течение всей взрослой жизни. Поддержание соответствующего восприятия запахов со временем нуждается, чтобы каждый новый нейрон проецировал аксон в соотв. место в обонятельной луковице. Эксперименты, в которых нейроны. экспрессирующие определенный рецептор, устранялись вплоть до позднего постнатального развития, показали, что способность организовывать гломерулы с помощью аксонов первичных обонятельных нейронов сохраняется даже у взрослых. Organization within the Olfactory Bulb Принцыпы, которые организуют соединения внутри обонятельных луковиц у млекопитающих, остаются предметом интенсивных исследований. Существование зон или доменов экспрессии рецепторов в обонятельном эпителии создаёт грубую топографическую организацию, в которой дорсально-экспрессируемые рецепторы проецируются в дорсальные регионы ОВ. Более того, высоко гомологичные рецепторы проецируются в гломерулы, которые находятся в тесном соприкосновении в ОВ (Strotmann et al., 2000). Подобное распределение согласуется с присутствием кластрированных реакций на запах в отдельных регионах луковиц. Всё это указывает на то, что механизм центр-окружения, подобный наблюдаемому процессингу в сетчатке. м. использоваться для заострения профиля реакции исходящих нейронов с этой системе. Недавно было продемонстрировано, что т. наз. "short axon" клетки , которые окружают центры процессинга гломерул, посылают аксоны прочь в более чем 20 гломерул, чтобы обеспечить механизм центр-окружение (Aungst et al. 2003) (Рис. 2). 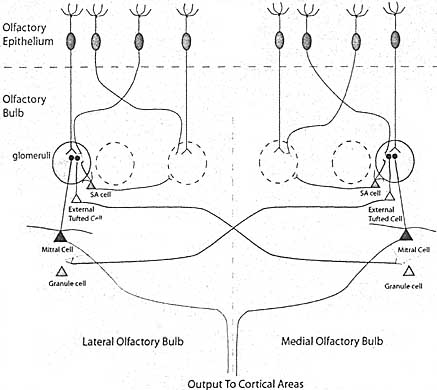 Figure 2. Schematic of Connections in the Olfactory Bulb Figure 2. Schematic of Connections in the Olfactory BulbIndividual olfactory neurons are initially distributed in the olfactory epithelium. Axons on cells expressing the same receptor (green or orange) fasiculate before reaching their target glomeruli. Mitral cells, the major output neurons of the olfactory bulb, and external tufted cells that project between mirror-symmetric medial and lateral gtomeruli receiving input from the same receptor are shown. The short axon (SA cells) that mediate center-surround refinement of olfactory signals are indicated in blue. Other major neuronal cells that surround the glomeruli and axonal projections of some cells are eliminated for clarity. Помимо этого существуют соединения на значительные расстояния, которые вносят вклад в процессинг сигналов в этой системе. Одним из примеров таких соединений м. служить генетически идентифицированные гломерулы, получающие проекции от клеток, экспрессирующих такой же OR ген (Belluscio et al., 2002). Один класс нейронов второго порядка в луковицах, наружные tufted клетки (Рис. 2), имеет дендритные разветвления в одном меченном гломеруле и распространяет свой аксон поперек луковицы в соотв. генетически меченный гломерул на медиальной стороне. Набор tufted клеток из меченных медиальных гломерулов устанавливает сходные реципрокные соединения с соотвествующими гломерулми латеральной поверхности. Хотя роль этих соединений не установлена, но, по-видимому, они совершенствуют избирательность реакции и снижают шумы в системе. The Next Frontier - Organization of Olfactory Аксоны mitral клеток , которые выходят из обонятельных луковиц проецируются во множественные области обонятельного кортекса, каждая из которых обладает самостоятельной функцией. Эти аксоны м. нести закодированную информацию, которую высшие разделы головного мозга интерпретируют как определенный запах и его интенсивность. Сложность стимулов, которые закодированы и нашей способности идентифицировать смеси и вычленять их составные, указывает на то, что д. происходить сложный процессинг в этих кортикальных областях. Необходимо знание распространение сигналов второго порядка нейронами к их соотв. мишеням, организации и логического обонятельного кодирования в этих высших областях. OR гены, нагруженные меткой. которая м. передаваться транс-синаптически нейронам второго порядка, выявили паттерн проекции для клеток, воспринимающих иннервацию от нейронов. экспрессирующих один OR (Zou et al., 2001). Эти исследования показали. что кластеры кортикальных клеток получают перекрывающиеся сигналы от множественных ORs. Развитие дополнительных более мощных методов для визуализации кортикальных мишеней для специфических субнаборов соотв. нейронов, необходимо для картирования этих путей и выяснения подлежащих механизмов обонятельного кодирования. Conclusion
White a complete understanding of olfactory perception that fascinated Lewis Thomas has not yet been achieved, the integration of molecular, physiological, and anatomical approaches has peeled away many mysteries in this sensory system. We are beginning to appreciate the centra) role of the olfactory receptors in defining the responses of individual cells and the organization of the peripheral and central processing circuitry. The challenges that await olfactory researchers, elucidating the processes utilized by the brain to read the patterns present in the olfactory bulb and develop a perception of simple and complex smells, are far more complex. The achievement of this goal, requiring creativity and interdisciplinary approaches, may finally reveal the inner workings of this wondrous sensory system and the brain itself.
| |

 → | K
→ | K