Многочисленные эксперименты, проведенные в лаб. P. Sengel’s показали, что замещение кусочка соматической мезодермы с одного уровня на др. вдоль cephalo-caudal оси ведет к развитию части спинального pteryla в соответствии с происхождением трансплантата. Не сегментированная соматическая мезодерма является т.о. уже регионально специфицированной (Mauger, 1972a). Dr. Kieny and Dr. Weydert безуспешно пытались найти различия в мРНК, присутствующих в разных сомитах. Сегодня известно, что регионализация сомитов базируется на экспрессии различных наборов Hox генов и что эти транскрипционные факторы присутствуют в небольших количествах (among others: Duboule, 1992; Duboule and Dolle, 1989). Напр., cHoxC-8 участвует в спецификации соматических клеток, которые дают дорсальный торакальный дермис с помощью E6 (Kanzler et al ., 1997).
Как же осуществляется спецификация дермальных предшественников из дермомиотома и соматоплевры? Интересен факт позитивной роли нервной трубки не только в формировании дорсального pteryla (Mauger, 1972b), что понятно благодаря близости этих двух структур, но и также в инициации дополнительных pteryla из midventral apterium (Sengel and Kieny, 1967a and b). Во многих лаб. идентифицированы разные типы диффузных молекул, синтезируемых в нервной трубке. Благодаря этому были решены молекулярные основы спецификации дорсальных и вентральных pterylae в моей лаб. (Olivera-Martinez et al ., 2001 and 2002; Fliniaux et al ., submitted). Более того, механизмы образования перья-образующего дермиса, по-видимому, отличаются на дорсальной и вентральной сторонах эмбрионов кур, не обязательно за счёт разного типа участвующих молекул, но благодаря их роли, времени их экспрессии и участии в сигнальных каскадах. Теперь мы лучше понимаем, почему участки glabrous кожи получаются после трансплантации кусочка презумптивной территории midventral apterium на место сомита (Mauger 1972a). В противоположность ранней интерпретации, сегдня изсестно, что соматоплевра даже в своей наиболее дистальной части всё ещё поддатлива (malleable) в E2/E3 (Fliniaux et al ., 2004). Однако. когда этот кусочек соматоплевры трансплантируется вблизи нервной трубки, то он не получает необходимых сигналов в соотв. время. Так становится понятным образование дополнительных pteryla из соматоплевры области midventral apterium, и даже из внеэмбриональной соматоплевры. Недавние результаты, полученные в моей лаб. (Fliniaux et al ., 2004), показывают, что молекулярный сигнал, который возникает в спланхноплевре, фактически неважен для увеличения плотности мезодермальных клеток. Этот факт уже отмечался, но ранее это объясняли с механической точки зрения. Более того формирование перьев внкэмбриональной соматоплеврой демонстрирует также, что морфогенез перьев является базовой программой эктодермы птиц. Морфогенез чешуек требует некоторых дополнительных модификаций в основном для предупреждения их роста (Kanzler et
al ., 1997, and see Prin et al ., 2004).
Наконец, большинство экспериментов проведено на эмбрионах кур. Поэтому определение точного источника и миграторного поведения сомитного или splanchnopleural происхождения дермы всё ещё остаётся открытым вопросом у мышей.
Сайт создан в системе
uCoz ISABEL OLIVERA-MARTINEZ, INGRID FLINIAUX, SYLVAIN MISSIER,
JEAN P. VIALLET and JACQUES THELU
ISABEL OLIVERA-MARTINEZ, INGRID FLINIAUX, SYLVAIN MISSIER,
JEAN P. VIALLET and JACQUES THELU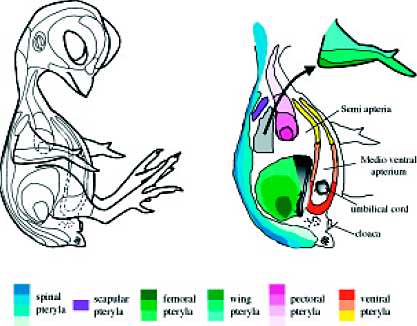 The different chick skin pterylae. Modified from Mayerson and Fallon (1985). Diagram by I. Fliniaux.
The different chick skin pterylae. Modified from Mayerson and Fallon (1985). Diagram by I. Fliniaux.  Right). Dorsal (A) and ventral (B) skin of 11-day chick embryo. Abbreviations: cp, cervical part of the spinal pteryla; dfp, dorsal part of femoral
pteryla; mva, medioventral apterium; pp, pectoral pteryla; sa, semiapterium; scp, scapular pteryla; spp, spinal pteryla; u, umbilical cord; vfp, ventral
part of the femoral pteryla; vp, ventral pteryla. Dissections and photographs by A. Mauger (A) and I. Fliniaux (B).
Right). Dorsal (A) and ventral (B) skin of 11-day chick embryo. Abbreviations: cp, cervical part of the spinal pteryla; dfp, dorsal part of femoral
pteryla; mva, medioventral apterium; pp, pectoral pteryla; sa, semiapterium; scp, scapular pteryla; spp, spinal pteryla; u, umbilical cord; vfp, ventral
part of the femoral pteryla; vp, ventral pteryla. Dissections and photographs by A. Mauger (A) and I. Fliniaux (B).