Ottawa naked (OT) аутосомно рецессивная мутация, обусловливающая практически полное отсутствие перьев у вылупляющихся цыплят. Частое webbing пальцах II и III ассоциирует в голой кожей. Более того, эмбрионы, исследованные на 6-й день, имеют часто аномалии нервной трубки или даже полное отсутствие хвостовой области. Цыплята редко достигают взрослой стадии. Если они выживают, то у взрослых м. сформироваться немного down перьев, но большая часть тела совершенно голая. На E6.75 (ст. HH 30) формирование дермиса очень нерегулярное в дорсальной области и по большей части субэктодремальная мезенхима остаётся рыхлой (Fig. 1A). На E8.5 иногда появляются зачатки немногих перьев, правда, с задержкой на 2 дня по сравнению с нормой. На E10 (Fig. 1B), если формируются зачатки немногих перьев, то их количество, диаметр и локализация в дорсальном поле (macropattern) и их расположение (micropattern) очень изменчивы, но почти симметричны на каждой стороне от срединной дорсальной линии. Плотный дермис отсутствует в большинстве дорсальных областей (Fig. 1 C,D), хотя он формируется и в последствии даёт аномальных размеров конденсаты дермы (Fig. 1 E,F). Гетерогенетические дермально/эпидермальные рекомбинации были осуществлены на ст. HH 31 между мутантными эмбрионами (OT/OT) и диким типом (WT/WT) и затем трансплантированы на 6 дней на хорионаллантоисную мембрану кур. Каждому мутантному эмбриону проводили три трансплантата: контрольный; OT/OT dermis/WT/WT epidermis; и WT/WT dermis/OT/OT epidermis. Во всех случаях (n=6) получены одинаковые результаты. Контрольные и с мутантным дермисом рекомбинанты не имели перьев или давали немного перьев. В тех редких случаях образования немногих перьев (Fig. 1 G,H), их образование задерживалось на 2-3 дня по сравнению с рекомбинантами с дермисом дикого типа. В последнем случае дермис давал большие количества перьев (Fig. 1I), и соотв. зачатки дифференцировались быстро спустя день после рекомбинации. Ottawa naked дефект, т.о., затрагивает формирование плотного дермиса, тогда как Ottawa naked эпидермис функционирует нормально. Образование дермиса, однако, нарушено независимо от происхождения предшественников дермы, т.е. нейрального гребня (Couly and Le Douarin, 1988) для дермы лица, дермомиотома (Mauger, 1972) для дорсальной дермы или соматоплевральной мезодермы (Christ et
al ., 1983) для вентральной и кожи ног. Хотя молекулярные механизмы, которые обусловливают спецификацию предшественников дермы, по-видимому, отличаются в областях, напр., спины (Olivera-Martinez et al ., 2000, 2001, 2002 and 2004) и живота (Fliniaux et al ., 2004). Более того, как только сформируется субэктодермальная мезенхима, то мы м. подозревать, что затрагивается только densification дермиса. Это указывает на то, что или мезенхимные клетки затрудняют их пролиферацию/миграцию вблизи к эктдермы или что они не воспринимают специфические сигналы от эктодермы или что сигналы на недостаточном уровне.
When the dense dermis is not redistributed to
form dermal condensation: the chick Scaleless mutant
Scaleless является аутосомно рецессивной мутацией, которая не охарактеризована ещё генетически, но явилась предметом многих исследований. Scaleless цыплята имеют в основном гладкую кожу, в основном свободную от перьев, а их tarsometatarsus и ноги лишены чешуек. Разбросанные перья присутствуют на голове, бёдрах, бедренных и хвостовых pterylae (Fig. 2A). Яйца от гомозиготных SC/SC мутантов получены J.L. Pierro (Center for Environmental
Health, University of Connecticut, Storrs). Гистологические наблюдения эмбрионов на ст. HH 30 показали, что дорсальный дермис формируется нормально и представлен плотным дермисом, покрывающим рыхлый дермис (Fig. 2B) (Viallet et al ., 1998). T. о., scaleless эмбрионы (Viallet et al .,1998; Dhouailly et al ., 1998; Widelitz et al., 2000; M. Harris, personal communication), как и эмбрионы дикого типа (Wessells, 1965; Dhouailly et al ., 2004), формируют дермис в областях, которые соответствуют pterylae, и которые характеризуются повышенной плотностью фибробластов. У scaleless эмбрионов, однако, не происходит следующей ступени. Образование перьев нуждается в сегрегации, по крайней мере, двух типов дермальных клеток, благодаря перераспределению клеточной популяции, которая формирует плотный дермис (F. Michon, unpublished data). Первый тип д. формировать дермальные конденсаты и д. затем обеспечиваться морфогенетическими свойствами и участвовать в формировании перьев. Второй тип д. формировать ординарный рыхлый дермис, лежащий под эпидермисом, не образующим перьев. DiI эксперименты (Jiang et
al ., 1999) показали, что две судьбы дермальных фибробластов всё ещё м.б. передетерминированы на ст. E8 (HH 33), т. e. когда уже сформированы дермальные конденсаты. Notch путь, как известно, играет роль в бинарном выборе (Artavanis-Tsakonas et al ., 1995; Simpson, 1997), а при гибридизации in situ эмбрионы кур дикого типа обнаруживают, что Delta-1 транскрипты гетерогенно распределяются в формируемых дорсальных дермальных конденсатах на E7/E8
(Viallet et al ., 1998). Напротив у scaleless эмбрионов той же стадии Delta-1 транскрипты гомогенно распределены в плотном дермисе (Fig. 2C) (Viallet et al ., 1998). Более того, избыточная экспрессия Delta-1 у эмбрионов дикого типа ведет к образованию больших, эктопических вторичных apteria (Viallet et al .,
1998). Goetinck and Abbott, 1963; Sengel and Abbott, 1963 показали, что scaleless дефект экспрессируется посредством эпидермиса, т.к. scaleless дермис функционирует нормально. Гетерохронные, гетерогенные рекомбинации продемонстрировали, что scaleless дермис наделен способностью индуцировать придатки на ранней стадии и быстро теряет ее из-за отсутствия взаимодействия с эпидермисом дикого типа (Dhouailly and Sawyer, 1984; Song and
Sawyer, 1996). Гетерогенные рекомбинации scaleless дермиса с дикого типа эпидермисом ведут после 36 ч к преобразованию паттерна экспрессии Delta-1 в дермисе (Fig. 2D), это сопровождается спустя 6 дней культивирования на хорионаллантоисной мембране появлением перьев (Fig. 2E) (Viallet et al ., 1998). Обратные рекомбинанты дорсального scaleless эпдермиса с дикого типа дермисом ведут к гомогенному распределению транскриптов Delta-1 в поверхностном дермисе спустя 36 ч. (Fig. 2F), а 6 днями позднее к появлению гладкого экспланта (Fig. 2G) (Viallet et al ., 1998). Song et al ., 1996 показали, что дефект у scaleless эмбрионов заключается в отсутствии экспрессии FGF4 эпидермисом. Тогда же наша группа проделала сходного типа эксперимент, но кусочки дорсальной scaleless эмбриональной кожи покрывались шариками с FGF2 и трансплантировались на хорионалантоисныую мембрану. Появлялись прекрасно сформированные перья (Viallet et al ., 1998), более того, зачатки перьев возникали последовательно вблизи прикрепленных шариков, указывая, что эти шарики не замещают эпидермальные плакоды, как это предположили Song et al . (1996), а вместо этого сообщают и распространяют общее необходимое сообщение дермису. Эндогенная передача сигналов FGF4 от эпидермиса в дермис у эмбрионов дикого типа является т.о. пермиссивным сигналом, который м. непосредственно взаимодействовать с экспрессией Delta-1, делая возможным (Viallet et al., 1998;
Dhouailly et al., 1998) формирование дермальных конденсатов в перья- и чешуйки-формирующих областях. Дермальные конденсаты для чешуек, также как и формирование плакод, которые предшествуют организации плотного дермиса, чётко различимы на целых участках дермиса и эпидермиса после их разделения (Dhouailly, 1984). FGF4 не составляет "feather-message", как было предположено третьей группой (Widelitz et al ., 1996). Они получили избыточное образование перьев в подкрыловом semiapterium при использовании шариков FGF2, мы не работали с midventral apterium (Dhouailly et al ., 1998). Различия между верхним дермисом semi-apterium и тем, что в apterium в том, что первый обычно формирует плотный дермис, хотя и с задержкой, а также немного случайно распределенных перьев, тогда как последний остаётся рыхлым и полностью голым (Sengel et al ., 1969; Sengel, 1976). Более того шарики FGF2 позволяют scaleless коже ног формировать scutate чешуйки (Dhouailly et al ., 1998; Prin and Dhouailly, 2004).
The first dermal induction and the initiation/patterning
of appendages
Гетероспецифические рекомбинации между дермисом и эпидермисом осуществлены 30 лет назад с использованием тканей кожи от эмбрионов ящериц, кур и мышей (Dhouailly, 1973, 1975, 1977). Все они дали сходные результаты. Специфичность кожных придатков, т.е. образование зачатков чешуек, перьев или волос зависит от эпидермиса, тогда как их инициация, формирование паттерна и расположение зависит от дермы. Дермис передаёт свои инициальные запускающие сигналы, чтобы инициировать морфогенез придатков, а также специфицирует их размер и паттерна распределения, а эпидермис отвечает в соответствии со своим генетическим потенциалом. Эти зачатки не могут, однако, дать зрелые придатки. Напр., волосы формирующий дермис мышей индуцирует большие количества двухразмерных классов зачатков чешуек в эпидермисе ящериц (Fig. 3A) и зачатков перьев в эпидермисе кур, соответствующих центральным большим и латеральным малым первичным фолликулам шкуры мышей (Fig. 3B). Скорее всего мышиный вибрисы-формирующий дермис индуцирует небольшие количества гигантских зачатков чешуек в эпидермисе ящериц (Fig. 3C) и гигантские зачатки перьев в эпидермисе кур в соответствии с паттерном верхней губы у мышей (Fig. 3D). Ряд сигнальных факторов, как было установлено, экспрессируется в плакодах, а также в дермальных конденсатах. Bmp 2, 4 and 7 и follistatin (Chuong, 1998) экспрессируются в зачатках перьев во время формирования паттерна, тогда как только gremlin экспрессируется в дермисе между зачатками (Bardot et al ., 2004). Эктопическая экспрессия Bmp2 и 4 ведет к ингибированию зачатков перьев и реципрокно избыточная экспрессия noggin, follistatin и gremlin индуцирует образование эктопических или увеличенных перьев (Noramly and Morgan, 1998; Patel et al ., 1999). Интересно, что все активаторы перьев (BMP антагонисты, FGF, Shh) и большинство ингибиторов (BMP2, 4) экспрессируются в примордиях. Это ведет к предположению о диффузии репрессорных\активирующих сигналов (Chuong, 1998).
Динамическое формирование паттерна эмбриональной дорсальной кожей кур м. сопровождаться обнаружением транскриптов BMP-2 (Noramly and Morgan, 1998) сначала в формирующихся плакодах, затем в дермальных конденсатах. Вся дорсальная lumbar кожа восстанавливается у эмбрионов кур ст. HH 31 , а экспланты, культивируемые in vitro дают урожай спустя 8, 12 и 18 ч. Во время 0 (HH31) экспрессия BMP-2 в клетках плакод делает возможной идентификацию 5/7 отдельных рядов зачатков перьев (Fig. 4A). Спустя 8 ч. культуры экспрессия BMP-2 обнаруживается с трудом (Fig. 4B) и не обнаруживается спустя 10 ч, если имеется полная потеря периодического паттерна. Спустя 12 ч, этот паттерн восстанавливается повторно с некоторыми модификациями, такими как слияния, которые происходят вдоль линий шевронов (chevrons) (Fig. 4C). Необходимо отметить затем в дорсальном pteryla, что микропаттерн зачатков перьев в основном гексагональный, так что каждое перо окружено 6 др. перьями. расстояния между перьями крупнее в продольном направлении, чем в боковом, так что перья выглядят аранжированными в виде косых поперечных рядов или шевронов. В туловище шевроны открыты в направлении хвоста (Sengel, 1976; Dhouailly et al ., 2004). Спустя 18 ч, почти весь кожный эксплант образует паттерн de novo (Fig. 4D),правда с некоторыми аномалиями, такими как латеральные слияния и гетероогенность в диаметре зачатков перьев. Эти эксперименты (F. Michon, unpublished data) показали, что кожа обладает способностью автономно перестраивать новый периодический паттерн, если он был нарушен механическими стрессами из-за выделения и органотипического культивирования. Необходимо отметить в этом эксперименте, что аксиальный ряд возобновляется первым. Однако, присутствие кожи, соответствующей инициальному первичному ряду, не обязательно для повторного формирования паттерна интегумента: появляется новый доминантный ряд, латеральнее по отношению к инициальному (Novel, 1973). Появлению эпидермальных плакод предшествует организация плотного дермиса в дермальные конденсаты (Sengel and Rusaцuen, 1968; Dhouailly, 1984) и мы м. предположить, что первая дермальная индукция инициирует формирование плакод, которые в свою очередь делают возможной и направляют миграцию клеток в дермисе, ведущей к перераспределению фибробластов плотного дермиса. Т.к. первые дермальные сообщения, ответственное за инициацию и формирование паттерна придатков, остаются невыясненным у этих трёх классов амниот, то было предположено, (Dhouailly, 1977), что они обеспечиваются одним и тем же типом молекул, эволюционно хорошо законсервированных. Позднее это действительно было показано, для эмбрионов кур и мышей (Chuong, 1998). Что эти сигналы принадлежат семейству Wnt было подтверждено в экспериментах с ингибитором Wnt Dickkopf-1 (Dkk-1). Dkk-1 является растворимым ингибитором Wnt, который ингибирует активацию Wnt рецепторов Frizzled путём соединения с ко-рецептором low-density-lipoprotein-receptor-related protein 6 (LRP6)(Semenov et al ., 2001; He et al ., 2004). Когда диссоциированные дермальные клетки эмбрионов кур были трансдуцированы RCAS- Dkk-1 и затем реагрегированы и покрыты эпидермисом, то формирование перьев ингибировалось (Chang et al ., 2004). Dkk, однако, не делает отличий между разными Wnts, а специфические антагонисты для каждого из членов этого большого семейства ещё не установлены.
The second dermal induction and feather organogenesis
Ассоциация эпидермиса кур с формирующим волосы дермисом мышей продуцирует арестованные зачатки перьев, которые распознаются по аномальным гребням зубцов (Dhouailly, 1973). ОНи образуются в результате складывания в стек клеток, соответствующих будущим зубцам и barbule клеткам. Несмотря на то, что цитодифференцировка клеток перьев не зависит от дермальных сигналов, если образование пера было запущено первыми дермальными сообщениями, исходящими из мышечного дермиса, роста и типичной архитектурной организации перьевых филамент не происходило.
У кур количество зубцов (barbs) в neoptile перьев варьирует от 11
до 16, а остов (rachis), формируемый за счёт слияния передних зубцов, рудиментарен. Напротив количество зубцов в neoptile перьев уток варьирует от 18 до 26 и передний rachis хорошо развит. Более того, teleoptile remiges появляется вскоре после рождения у кур, но позднее у уток. Количество barb ridges в эмбриональных филаментах перьев, которое соответствует таковому количеству barbs в neoptile пера; их частичное слияние в основании пера, чтобы сформировать rachis; время образования второй генерации, ювенильных перьев и их форма, являются видоспецифичными характеристиками, которые все управляются вторичными дермальными сообщениями (Dhouailly, 1970). Эксперименты по гетероспецифическим рекомбинациям зачатков крыловой эктодермы и мезодермы между курами и утками дали чёткие результаты. Морфологические характеристики перьев детерминируются дермисом за исключением количества и формы barbule клеток, это подтверждает видовое происхождение эпидермиса (Fig. 5 A-C).
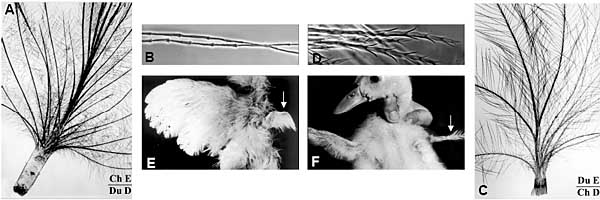 Fig. 5. Chimeric neoptile feathers produced by heterospecic duck/chick forelimb ectoderm/ mesodermal pulp recombinants. The dermis is responsible for the feather architecture and the time of replacement of the neoptile by the teleoptile feather, while the shape of epidermal barbule cells
is conferred by the epidermal species.(A) A typical duck-type neoptile feather obtained from the association of chick epidermis and duck dermis. Note
the formation of 26 barbs, most of them being attached to a well developed rachis. (B) Detail of the barbules of the feather shown in (A), showing a typical chick-type morphology, i.e. a succession of cylindrical cells, slightly swollen at their distal tip. (C) Typical chick-type neoptile feather obtained from the association of chick dermis and duck epidermis. Note the formation of 12 barbs. (D) Detail of the barbules of the feather shown in (C), showing a typical duck-type morphology, i.e. a succession of cylindrical cells, with two spiny protrusions at their distal tip. (E) A chick host, two weeks old, bearing a chimeric right wing composed of a duck mesenchymal pulp associated with a chick ectoderm. The chimeric wing is perfectly developed, but is covered only with neoptile duck-type feathers, while the left host wing is covered by the second generation, the juvenile teleoptile feathers. (F) A duck host, two weeks
old, bearing a chimeric right wing composed of a chick mesenchymal pulp associated to a duck ectoderm. The chimeric wing is in this case poorly
developed, because the microsurgery was not perfect, but in the second generation, the juvenile teleoptile chick-type feathers had formed (Dhouailly,
1970).
Fig. 5. Chimeric neoptile feathers produced by heterospecic duck/chick forelimb ectoderm/ mesodermal pulp recombinants. The dermis is responsible for the feather architecture and the time of replacement of the neoptile by the teleoptile feather, while the shape of epidermal barbule cells
is conferred by the epidermal species.(A) A typical duck-type neoptile feather obtained from the association of chick epidermis and duck dermis. Note
the formation of 26 barbs, most of them being attached to a well developed rachis. (B) Detail of the barbules of the feather shown in (A), showing a typical chick-type morphology, i.e. a succession of cylindrical cells, slightly swollen at their distal tip. (C) Typical chick-type neoptile feather obtained from the association of chick dermis and duck epidermis. Note the formation of 12 barbs. (D) Detail of the barbules of the feather shown in (C), showing a typical duck-type morphology, i.e. a succession of cylindrical cells, with two spiny protrusions at their distal tip. (E) A chick host, two weeks old, bearing a chimeric right wing composed of a duck mesenchymal pulp associated with a chick ectoderm. The chimeric wing is perfectly developed, but is covered only with neoptile duck-type feathers, while the left host wing is covered by the second generation, the juvenile teleoptile feathers. (F) A duck host, two weeks
old, bearing a chimeric right wing composed of a chick mesenchymal pulp associated to a duck ectoderm. The chimeric wing is in this case poorly
developed, because the microsurgery was not perfect, but in the second generation, the juvenile teleoptile chick-type feathers had formed (Dhouailly,
1970).
Более того, время обновления придатков также зависит от дермиса (Fig. 5 E,F). Соотв. дермально/эпидермальные взаимодействия д. происходить в фолликуле пера, между дермальным сосочком и кольцом (стенкой) недифференцированных эпидермальных клеток. Последние пролиферируют и подразделяются на barb ridges, которые сливаются с передней частью кольца, чтобы дать rachis, с дополнительными barb ridges, образующимися в задней части кольца. Формирование rachis, также как и добавление новых barb ridges, наиболее выражено в неоптиле пера у уток, чем у кур и осуществляется посредством общего роста teleoptile пера у обоих видов.
Несколько сигнальных факторов экспрессируется в развивающихся перьях, включая BMP2, BMP4, Noggin и Shh (Yu et al ., 2002; and 2004). BMP4 экспрессируется в основном в дермальных сосочках, тогда как Noggin экспрессируется в его задней области, где начинают формироваться barb ridges.
Shh экспрессируется во внутреннем слое и является существенным для индукции апоптоза, а , следовательно, расщепления barb ridges. Группа Chuong в серии элегантных экспериментов показала, что с помощью добавления RCAS вирусов, несущих эти гены или доминантно негативные гены, в фолликулы выщипанных (plucked) teleoptile перьев, то во время регенерации формируются аномальные перья. Избыточная экспрессия Noggin, антагониста BMP, увеличивает количество barb и даже вызывает расщепление, тогда как избыточная экспрессия BMP2 и BMP4 вызывает образование гигантских rachis и слияния barb. Супрессия Shh ведет к формированию остаточной мембраны между barbs. Поэтому очень возможно, что существуют различия в уровне экспрессии Noggin и BMP4/2 дермальным сосочком фолликула neoptile пера у кур и уток.
Conclusion: the molecular events which mediate dermal/
epidermal interactions
Молекулярные события, которые лежат в основе клеточных взаимодействий во время морфогенеза кожи т.о., частично задокументированы в результате изучения паттернов экспрессии и эктопических воздействий. Большинство недавно описанных онтогенетических сигнальных путей участвует в одном или всех этих взаимодействиях (суммировано на figure 6).
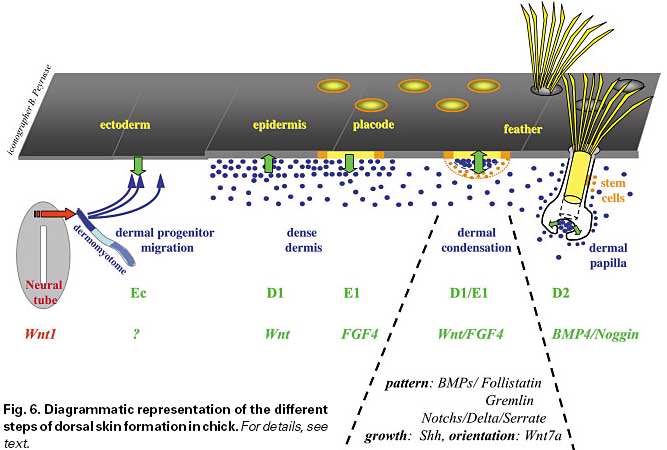
Среди этих сигналов путь Wnt ассоциирован прежде всего с формированием дорсального дермиса. Wnt1 из дорсальной части нервной трубки индуцирует спецификацию дермальных предшественников из дорсальной части дермомиотома (Olivera-Martinez et al ., 2001, 2002 and 2004). Дермальные предшественники экспрессируют Wnt 11, который м. участвовать в их миграции в субэктодермальное пространство (Olivera-
Martinez et al ., 2002, 2004). Wnt 1, 3a и 5a в зачатках перьев и Wnt 11 в области дермиса между зачатками контролируют форму зачатков перьев, тогда как Wnt7a участвует в передне-задней ориентации во время роста зачатков перьев (Chang et al ., 2004; Widelitz et al ., 1999; Chuong et al ., 1996). На ранней стадии уровень β-catenin гомогенен по всему эпидермису поля пера и затем ограничивается плакодой (Widelitz et al ., 2000). Мы полагаем, что эта генеральная экспрессия β-catenin является реакцией эпидермиса на генеральные сигналы Wnt, исходящие от дермы. Затем эти сообщения оказываются ограниченными примордиями у эмбрионов дикого типа, тогда как они остаются размазаны по всему тракту полей у scaleless эмбрионов (Widelitz et al ., 2000). Во время формирования паттерна окрашивание ядерного β-catenin увеличивается в плакодах и теряется в эктодерме, которая воспринимает меж-фолликулярную судьбу, кроме того, форсированная экспрессия β- catenin индуцирует образование эктопических перьев (Noramly et al ., 1999; Widelitz
et al ., 2000). Ограничение экспрессии β-catenin также как и экспрессии Delta-1 м.б. следствием экспрессии FGF4 в эпидермисе (Viallet et al ., 1998; Song et al., 1996). Путь Notch м. служить для стабилизации паттерна зачатков перьев (Viallet et al ., 1998). Сoupled BMP4/BMP антагонист выявляется первым во время спецификации поля пера на животе (Fliniaux and Viallet unpublished data). Во время образования pteryla временная экспрессия BMP2 обнаруживается в эпидермисе, тогда как BMP антагонисты gremlin и follistatin экспрессируются в подлежащем дермисе и эпидермисе, соотв. (Noramly et al ., 1998; Bardot et al ., 2004; Patel et al ., 1999). Если происходит формирование паттерна, то BMP2, 4 и 7 и BMP антагонисты за исключением gremlin экспрессируются в зачатках. Это наблюдение ведет к модели, базирующейся на активации посредством дифференциальной диффузии активаторов и ингибиторов для формирования периодического паттерна (Chuong, 1998). Позднее на ступени органогенеза пера пара BMP/antagonist (в данном случае Noggin) также вовлекается в формирование паттерна barb ridges (Yu et al ., 2002, 2004). Какова роль Shh? Он экспрессируется 2 дня энтодермой кур (Watanabe et al ., 1998) и м. играть роль в ингибировании экспрессии BMP4 в somatopleure в животе и тем самым м. позволять формирование перьевого поля. На ранней стадии формирования паттерна Shh экспрессируется в плакодах, тогда как его рецептор Ptc экспрессируется и в плакода и в дермальных конденсатах (Ting-Berreth and Chuong, 1996; Jung et al ., 1998; Morgan et al., 1998). С нашей точки зрения, Shh участвует в активации клеточной пролиферации и дермиса и эпидермиса. Форсированная экспрессия Shh (или воздействие Shh) вызывает эктопическое формирование перьев в pteryla, semi-apteria и даже в midventral apterium (Fliniaux et al., 2004) и, по-видиому, существенно усиливает плотность дермиса над критическим порогом. Фактически, Shh строго экспрессируется в зачатках перьев, которые удлиняются в филаменты пера, и слабо экспрессируется в перекрывающихся scutate чешуйках и едва обнаружим и коротко живёт в reticulate чешуйках, которые не перекрываются (Prin et al ., 2004). Ингибирование передачи сигналов Shh у кур (Prin and Dhouailly, 2004) м. приводить к аресту роста перьев. Shh нокаутные мыши (Chiang et al ., 1999) имеют арестованные зачатки волос. Всё это ведет к заключению, что Shh является генеральным активатором роста кожных придатков. Более того, Shh также позднее участвует, делая возможным апоптоз внутреннего эпидермального слоя и тем самым расщепление barb ridges во время органогенеза пера (Yu et al ., 2002, 2004).
Однако, формирование плотного дермиса и особенно природа сигналов, исходящих от эктодермы, также как миграция дермальных клеток для формирования дермальных конденсатов ждут своего разрешения. Наша предварительная диаграмма (Fig. 6), м.б. закончена добавлением, какие транскрипционные факторы и какого типа внутритканевые взаимодействия запускают каждую из ступеней. Только тогда мы получим картину перья- и волосы-формирующего морфогенеза кожи, как это почти сделано для органогенеза зубов ( Pispa and Thesleff,2003).
Сайт создан в системе
uCoz