Craniofacial Development and the Evolution of the Vertebrates: the Old problems on a New Background  Zool. Sci. - 2005.- Vol. 22, № 1. - P. 1-19 | |
|
|
A brief history of craniofacial studies Существует ли архетип сегментов в черепе? Количество сегментов включенных в череп также нерешенный вопрос сравнительной эмбриологии (Goodrich, 1930; de Beer, 1937; Jarvik, 1980; Jefferies, 1986). "Проблема сегментации головы" впервые поставлена Goethe и его коллегами, полагавших, что череп позвоночных представляет собой ансамбль позвонков, обнаруживаемых в посткраниальной части туловища ('vertebral theory' черепа; Рис. 1; Goethe, 1790; rev. Gaupp, 1898; Owen, 1866; rev. Goodrixch, 1930; de Beer, 1937; Neal and Rand, 1946; Kurtani, 2--3).
В эру сравнительной эмбриологии, между 19 и 20 веком, вопрос о сегментации головы снова стал центральным. Это, по крайней мере частично, было простимулировано открытем 'head cavities' у эмбрионов акулы (Рис. 2; Gee, 1996; Kuratani, 2004a), появляющихся в виде мезодермальных coeloms (Balfour, 1878;), а также сегментного происхождения затылочных хрящей (см. ниже). Головные полости являются действительно источниками внешних глазных мышц (соматических-мышце-подобных скелетных мышц в туловище) и возникают они, по-видимому, сегментно, обычно в виде трех пар, каждая ассоциирована с одиночной pharingeal arch (PA), вентральным висцеральным элементом. Полости затем были обозначены спереди назад как premandibular, mandibular, hyoid полсости, иннервированные нервами oculomotor, trochlear и abducens, соотв., точно также как каждый из миотомов, иннервируется спинальными нервами по-сегментно (мфт Wijhe, 1882; rev. Goodrich, 1930; de Beer, 1937; Jarvik, 1980; Jefferies, 1986; Рис. 2). Т.о., голова позвоночных стала рассматриваться как набор сегментов, каждый состоящий из дорсальной соматической части и вентральной висцеральной части, точно также как туловище, состоящее дорсально из сегментированных сомитов и вентрально из несегментированной латеральной пластинки (источника висцеральных гладких мышц). 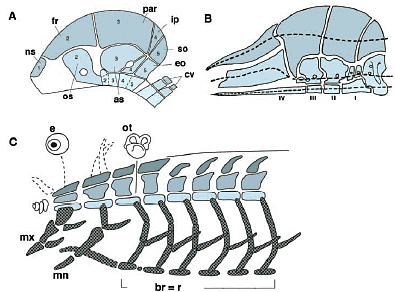  Череп также стал рассматриваться в терминах той же самой схемы, как продолжение сегментации головы. В целом череп позвоночных был подразделен дорсо-вентрально на нейрокраниум или капсулу, поддерживающую головной мозг и висцерокраниум, котороый поддерживает глотку (Gaupp, 1906; Goodrich, 1930; Gregory, 1933; de Beer, 1937; Portman, 1969; Рис. 3). Поскольку висцерокраниум сегментирован на единицы скелета фарингеальных (бранхиальных) дуг (напр., Sewerzoff, 1911), то было предположено, что и нейрокраниум также сегментирован, а pilar хрящи между корешками краниальных нервов часто приравнивались к нервным дужкам позвонков (Gaupp, 1906; de beer, 1937; Starck, 1980; rev. Kuratani, 2003). Т.о., череп позвоночных стал объясняться как имеющий один общий морфологический паттерн, вторично модифицированный специфичными для группы животных вариациями и дифференцировкой, так как это видно на специфичных для млекопитающих признаках, таких как косточки среднего уха, молоточек, наковальня, стремя, происходящие соотв. из articular, quadrate и hyomandibular соотв. у менее развитых животных.
Морфологическая схема свода черепа, описанная выше была выявлена прежде всего функционально, как поддерживающая ткань для головного мозга и глотки (Gregory, 1933; Рис. 3), хотя концепция гомологии не обязательно нуждается в сохранении исходных функций (Owen, 1866). Следовательно, передняя часть нейрокраниума, производное трабекулярных хрящей, как полагалось, представляют собой другую пару фарингеальных хрящей, принадлежащих к др. фарингеальной дуге, присутствовавшей у родоначальника (de Beer, 1931, 1937; см. также Kuratani et al., 1997, 2004). Эта гипотеза согласуется с теорией сегментации головы, которая рассматривает пре-мандибулярный сегмент, также как и тройничный нерв, как составной краниальный нерв; ophtalmic нерв, как предполагается, принадлежит к пре-мандибулярному сегменту, а mfxillomandibular нерв к мандибулярному сегменту (rev. Goodrich, 1930; de Beer, 1937; Jarvick, 1980).Т.о., ранние сравнительные исследования сконцентрировались на сведении каждого краниального сегмента к общей сегментной схеме, не задаваясь необходимыми вопросами о задействованных онтогенетических механизмах. Хотя онтогенетическое происхождение структур часто было фокусом дебатов, главным стержнем концепции была мезодермальная сегментация. Хотя вовлечение производной нервного гребня эктомезенхимы в краниогенезе подчеркивалось (Platt, 1893), это, по-видимому, не влияло на морфологическую схему черепа. 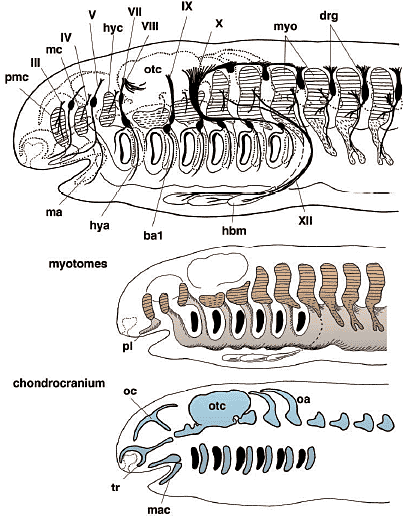  C успехами экспериментальной эмбриологии онтогенетическая роль производной нервного гребня эктомезенхимы в краниогенезе позвоночных и значение межтканевых взаимодействий оказалось принятым всеми (rev. Gans, Northcutt, 1983; Northcut, Gans, 1983; Hall, Horstadius, 1988; Le Dourin, 1982; Le Dourin, Kalcheim, 1999; Hall, 1999; см. также de Beer, 1926; Hanken, Hall, 1993). Однако, это не было правильно интегирировано с трансцендентальной морфологией до недавнего времени, когда молекулярная биология развития стала клеем, объединившим их.
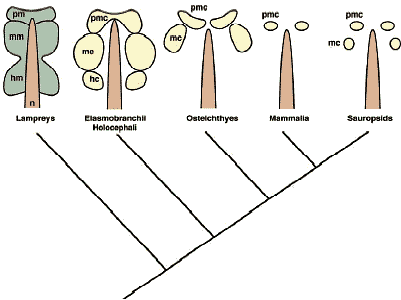
Базируясь на молекулярной генетике и экспериментальной эмбриологической технике современные исследования краниогенеза и эволюции позвоночных сконцентрировались на онтогенетических механизмах, участвующих в дифференцировке производной нервного гребня эктомезенхиме, регуляторных механизмах. лежащих в основе паттернов скоординированной экспрессии различных регуляторных генов, включая Hox и Dlx гены, и путях индуктивных сигналов, ведущих к дифференцировке специфических клеточных популяций. Segments in the mesoderm - somitomerism Preotic mesoderm Идея сегментации головы испытывала большее или меньшее влияние со стороны трансцендентальной или идеалистической философии классической морфологии, или ее производного сравнительной эмбриологии вплоть до начала двадцатого столетия. Те эмбриологи и анатомы верили, что даже если сегменты невидимы в своде черепа взрослого, сегментный материал д.б. видимым в зачатке краниума (rev. Goodrich, 1930; de Beer, 1937). Фактически, на заднем конце нейрокраниума (постотическая область) у большинства групп позвоночных затылочная кость развивается благодаря слиянию нескольких пост-отических производных сомитов, склеротомов. Это подтверждено современными экспериментами с использованием химер кур и перепелов (Couly et al., 1993), т.к. клетки перепела имеют уникальный ядерный маркер, отсутствующий в тканях кур. Пре-отическая область, с др. стороны, оказалась более проблематичной и предоставила строгие доказательства в подтверждение теории (мезодермальных)головных сегментов: открытие головных полостей у elasmobranch эмбрионов. Т.о., проблена сегментации головы м.б. подразделена на две части, пре-отическоую и пост-отическую в соотв. с проблемами, связанными с цефалической мезодермой и сомитами у эмбрионов, соотв. Preotic region - the head cavities Эволюционное и онтогенетическое значение головных полостей все ещё неясно (Рис. 2, 4, 5). Однако они м. и не быть примитивными свойствами для всех позвоночных, т.к. нет структур, сходных с головными полостями у эмбрионов миног или hagfish, наиболее базовой группы позвоночных, если головные полости определяются как настоящие целомные, выстланные тонким эпителием, то плавающая в голове мезенхима, которая состоит в основном из рыхлой соединительной ткани (фибробластов) (для мионг см. Kuratani et al., 1999; для классического описания Koltzoff, 1901; Damas, 1944). С др. стороны, головные полости были описаны у большинства представителей gnathostome таксона и они кажутся уменьшающимися в каудально-ростральном направлении вдоль кроны филогенетического древа (Рис. 5) . Следовательно, головные полости не кажутся представителями примитивных характеристик у позвоночных, хотя они м. представлять собой synapomorphy, по которой и определяются gnathostomes.   Онтогенетическая функция головных полостей всё ещё неизвестна, как и механизм формирования их паттерна. Концепция генеративных ограничений важна для сегментного паттерна в развитии: определенные паттерны, устанавливающиеся у ранних эмбрионов, м. влиять на последующее формирование паттерна рестриктивным консервативным способом, в результате возникает общий анатомический паттерн у разных групп животных. Типичным примером являются сомиты, которые формируют паттерн ганглиев дорсальных корешков и корешков спинальных нервов. Популяции клеток нервного гребня туловища и двигательные нервные волокна не являются инициально сегментированными, а являются вторичными подразделениями в присутствии сомитов, в результате возникает сегментированный паттерн спинальных нервов (Detwiler, 1934; Keynes, Stern, 1984; Tosney, 1988; re. Kuratani, 2003). Следовательно, первичным фактором в определенных паттернах явлдяется присутствие первичных сегментов. Представляют ли головные полости тот же самый сегментный паттерн, что и корешки краниальных нервов и ромбомеры? Ответ - нет. У эмбрионов акул, хотя полости и сохраняюдт топографические взаимоотношения с нервами в ходе всего развития, но положение корешков нервов сдвигается росто-каудально вдоль нейро-оси заднего мозга (Kuratani, Horigome, 2000). Более того, те же самые паттерны караниальных нервов появляются у большинства эмбрионов gnatostome с или без очевидными эпителиальными полостями. Т.о., полости, по-видимому, не функционируют как generative constraint (генеративные ограничители) в формировании паттерна периферических нервов. 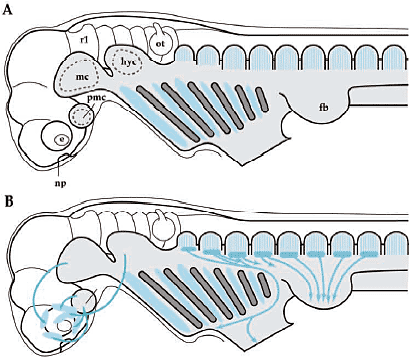 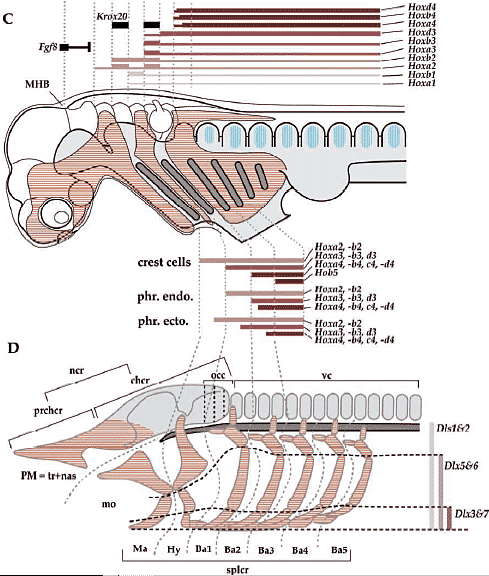  Somitomeres or regionalization of the head mesoderm Др. псевдосегментной структурой являются т.наз. "сомитомеры". В отличие от головных полостей, которые с самого начала эпителиальные, сомитомеры являются мезенхимными и лишены видимых сегментных границ. Настоящие сомитомеры первоначально наблюдали при сканирующей электронной микроскопии в виде неполных сегментных возвышений в параксиальной мезодерме области туловища до сомитогенеза. Сходные возвышения часто распознаются в цефалической мезодерме у некоторых видос позвоночных и сначала назывались "цефалическими сомитомерами" (Рис. 5). В целой серии работ было показано, что у всех видов позвоночных цефалическая мезодерма обнаруживает довольно консервативный топографический паттерн по отношению к эмбриональным структурам, таким как отическая плакода или оптический пузырёк, и обнаруживает стереотипические взаимоотношения с потоками клеток нервного гребня. В отличие от четкого анатомического паттерна головных полостей, присутствие сомитомеров, однако, проблематично. Согласно компьютерному анализу клеточных агрегатов не выявляется сегментного паттерна цефалических мезодермальных клеток (Freund et al., 1996). По крайней мере, сомитомеры не м.б. приравнены к головным полостям, которых насчитывается вполовину от сомитомеров. Поэтому возникают разные мнения относительно количества головных сегментов (Kuratani, 2004a).
Во-первых, имеются остатки сегментного паттерна в цефалической мезодерме, которые не создают сегментных ограничений др. структурами, подобно сомитам, создающим паттерн потоков клеток нервного гребня. Во-вторых, неясно, отражают ли псевдосегменты какую-либо онтогенетическую программу вообще.
Головная мезодерма не создает генеративных ограничений для какой-либо др. эмбриональной ткани, для создания краниального анатомического паттерна. Даже если сомитомеры представляют собой остатки сегментного паттерна, то морфологический паттерн головы позвоночных не сегментирован так, как в туловище. Ромбомеры и глоточные карманы скорее, чем цефалическая мезодерма, формируют паттерн краниальных нервов.
  В отношении второй проблемы имеется несколько причин опровергать врожденную программу сегментации цефалической мезодермы у позвоночных. Специфические мезодермальные области м.б. идентифицированы способом, сравнимым у разных видов, но не с помощью сегментации мезодермальной клеточной массы, а за счет регионализации мезодермы на несколько доменов в присутствии некоторых др. эмбриональных структур (Рис. 4А). Напр., мезодерма нижней челюсти м.б. предопределена как клеточная масса, обнаруживаемая в мандибулярной дуге (ограниченная сзади первым глоточным карманом), а гиоидная мезодерма ограничена каудально с помощью отической плакоды и второго глоточного кармана, а спереди первым глоточным карманом. После этой стадии регионализации цефалическая мезодерма выглядит как сегментно специфицирована, как это представляют сторонники сегментации, подобно Goodrich (1930; Рис. 6). Следовательно, даже если возможно морфологически различать специфические регионы в головной мезодерме способом. который удовлетворяет концепции морфологической гомологии среди разных видов, это еще не доказывает, что имеется сегментный паттерн в головной мезодерме, сходный с тем, что обнаруживается в постотических сомитах. 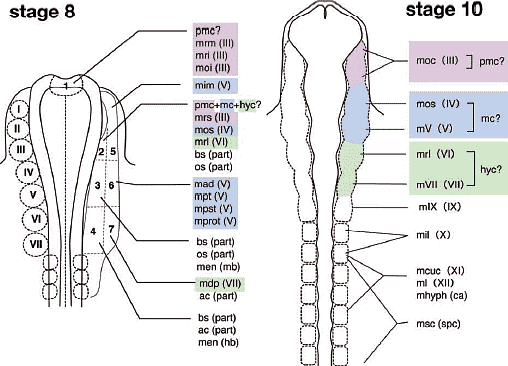 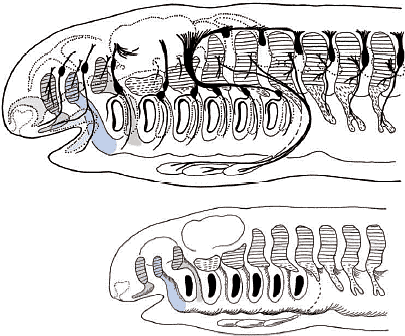  Из регионализованных мезодермальных "регионов", премандибулярная мезодерма, которая возникает относительно поздно в развитии из прехордальной пластинки и имеет четкую заднюю границу, м. представлять собой реальный "сегмент". При молекулярном анализе осцилляции экспрессии гена, связанного с сегментами, наблюдали только дважды у цефалической мезодерме эмбрионов кур, в премандибулярной мезодерме и в остальной мезодерме головы (Рис. 7; Jouve et al., 2002). Если сегментный компартмент предопределяется с помощью этих молекулярных функций, то этот результат подтверждает, что вся головная мезодерма позвоночных представлена одним большим сегментом, эквивалентным одиночному сомиту. Эта идея не согласуется с гипотетическим количеством мезодермальных сегментов, предполагаемых у родоначальника позвоночных (Holland, 2000), или сомитомеров. 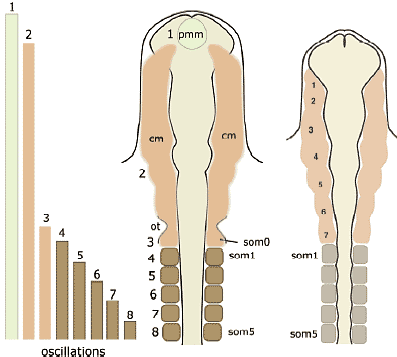 Fig.7. Somitomeres and somitogenetic genes. The morphological pattern of somitomeres in the chick embryo is shown on the right, as a simplified illustration. Hypothetical somitomeres are numbered. On the left is shown the oscillating expression of a somitogenetic gene, chairy, in the early chick embryo, based on Jouve wt al., (2002). Each oscillation is numbered together with the mesodermal part generated after that pscillation. Note that there are only two oscillations in the head mesoderm, one for the premandibular mesoderm, and the other for rest of the cephalic mesoderm. Fig.7. Somitomeres and somitogenetic genes. The morphological pattern of somitomeres in the chick embryo is shown on the right, as a simplified illustration. Hypothetical somitomeres are numbered. On the left is shown the oscillating expression of a somitogenetic gene, chairy, in the early chick embryo, based on Jouve wt al., (2002). Each oscillation is numbered together with the mesodermal part generated after that pscillation. Note that there are only two oscillations in the head mesoderm, one for the premandibular mesoderm, and the other for rest of the cephalic mesoderm. Transposition and homeotic transformation Как отмечалось выше, задняя часть нейрокраниума онтогенетически сегментирована ена сомиты (Рис. 3, 4). В сравнительной эмбриологии затылочная кость рассматривается как часть оригинального туловища, которая вторично ассимилирована и интегрирована в краниогенезе. Однако, количество окципитальных позвонков отличается у разных видов животных, указывая тем самым, что разные количества сегментов м. иметь одни и те же морфологические качественные особенности. Следовательно, морфологическая гомология скелетных элементов не м.б. уменьшена до серийного количества онтогенетических компартментов. Эта проблема д. решаться в терминах осевой спецификации позвоночного столба, с которой связаны паттерны скоординированной экспрессии гомеобокс-содержащих генов (Нох генов)(Рис. 8).
Нох гены кодируют транскрипционные факторы и ранжированы тандемно в ДНК, которая представляет Нох кластер. Т.к. в результате геномной дупликации имеется 4 Нох кластера в геноме у амниот и базовых позвоночных, хотя кластеры у teleost, по-видимому, подверглись дополнительному удвоению. Нох гены паралоги занимают эквивалентные позиции в кластерах и составляют paraloge group (PG) гены, которые нумеруются в направлении 3' -> 5', PG1-PG4.
В онтогенетической спецификации скелетных элементов, базирующейся на Нох генах, имеется тенденция т.наз. 'spatial colinearity', так что Нох гены, расположенные более 3' в кластере экспрессируются более кпереди, а те, что на 5' конце экспрессируются более кзади вдоль передне-задней оси туловища эмбриона (Рис. 8).
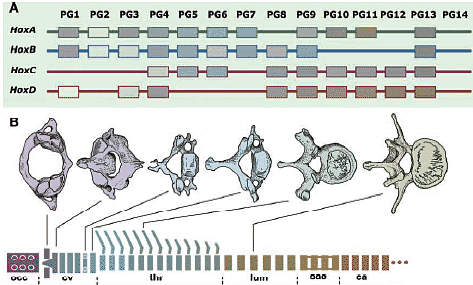 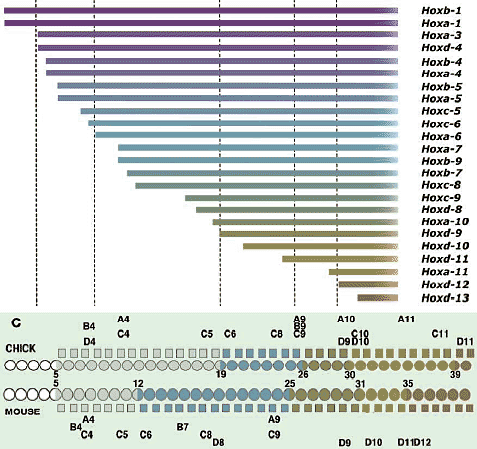 Fig. 8. Homeotic transsformation of the vertebral column and Hox genes/ (A) There are four Hox clusters (HoxA, to HoxD) in the genomes of non-teleost gnathostomes. (B) Schematic illustration of Hox-code-dependet vertebral specification in the mouse, based on Kessel (1992). Each Hox gene is expressed with a slightly different rostral expression boundary in the trunk, largely based on a nested, collinear pattern. Thereby, the provertebra at each level is specified by a specific set of Hox genes, facilitating the axial specification of the vertebral column. Note that the occipital bone, the posterior part of the neurocranium, is also specified as part of the vertebral column derived from an array of somites. (C) Comparison of the Hox codes of the chicken and mouse, based on Burke et al., (1995). Note that morphological identities are not associated with specific numbers of somites but with the expression domains of identical (homologous) Hox genes. ca, caudal vertebrae; cv, cervical vertebrae; lum, lumbar vertebrae; occ, occipital vertebrae; thr, thoracic vertebrae Fig. 8. Homeotic transsformation of the vertebral column and Hox genes/ (A) There are four Hox clusters (HoxA, to HoxD) in the genomes of non-teleost gnathostomes. (B) Schematic illustration of Hox-code-dependet vertebral specification in the mouse, based on Kessel (1992). Each Hox gene is expressed with a slightly different rostral expression boundary in the trunk, largely based on a nested, collinear pattern. Thereby, the provertebra at each level is specified by a specific set of Hox genes, facilitating the axial specification of the vertebral column. Note that the occipital bone, the posterior part of the neurocranium, is also specified as part of the vertebral column derived from an array of somites. (C) Comparison of the Hox codes of the chicken and mouse, based on Burke et al., (1995). Note that morphological identities are not associated with specific numbers of somites but with the expression domains of identical (homologous) Hox genes. ca, caudal vertebrae; cv, cervical vertebrae; lum, lumbar vertebrae; occ, occipital vertebrae; thr, thoracic vertebrae Т.к. эти гены обычно экспрессируются от определенного уровня оси и каудально, то каждый сомит вдоль оси д. экспрессировать специфический набор Нох транскриптов с гнездовым паттерном. такой паттерн экспрессии Нох генов наз. "Нох кодом" и функционирует как система обеспечивающая позиционные значения сомита на каждом уровне, так что он м. дифференцировать соотв. качественные особенности. Интересно, что одни и те же морфологические особенности позвонков, включая и окципитальные, кодируются гомологичными наборами Нох генов во всех группах животных, но не количеством сегментов. Следовательно, сомиты не добавляются и не теряются вторично во время эволюции или морфологические качественные особенности, ассоциированные с номерами сомитов. Однако, гетеротопический сдвиг в регуляции Нох генов (устанавливающего Нох код), по-видимому, м. служить основой эволюционных транспозиций качественных особенностей позвонков, как предполагает др. группа эмбриологов. Следовательно, регуляция Нох кода, по-видимому, меняется в ходе эволюции, создавая формулу позвонков, которые отличаются в каждой группе животных (Рим. 8В). Тот же самый механизм м. объяснить и эволюцию формулы позвонков (Burke et al., 1995; rev. Narita, Kuratani, 2005). 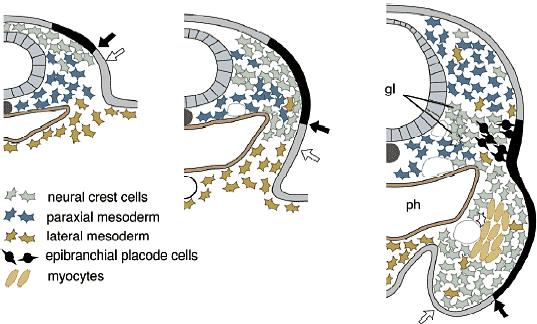 Fig.9. Development of cephalic crest cells. The distribution pattern of cephalic neural crest cells in the developing chiken embryo is diagrammatically illustrated, based on Noden (1988). Arising at the junction of the neuriectoderm and surface ectoderm, the crest cells in the head migrate ventrally and laterally along the superficial pathway, called the dirsolateral pathway. They finally occupy the PA, to form the ectomesenchyme surrounding the branchial arch muscle primordium derived from the cephalic mesoderm. Note that, at the latest stage, the crest-derived ectomesenchyme and mesoderm are dissociated dorsoventrally, and the future cranial base is composed of mesodermal cells lateral to the notochord. Ehite arrows indicate the leading edge of the migrating crest cells, and the black arrows the relative growth of the cephalic surface ectoderm. Note where the position of the epibranchial placode is mapped in the young embryo. gl, sensory ganglion of the branchial nerve; ph, pharynx Fig.9. Development of cephalic crest cells. The distribution pattern of cephalic neural crest cells in the developing chiken embryo is diagrammatically illustrated, based on Noden (1988). Arising at the junction of the neuriectoderm and surface ectoderm, the crest cells in the head migrate ventrally and laterally along the superficial pathway, called the dirsolateral pathway. They finally occupy the PA, to form the ectomesenchyme surrounding the branchial arch muscle primordium derived from the cephalic mesoderm. Note that, at the latest stage, the crest-derived ectomesenchyme and mesoderm are dissociated dorsoventrally, and the future cranial base is composed of mesodermal cells lateral to the notochord. Ehite arrows indicate the leading edge of the migrating crest cells, and the black arrows the relative growth of the cephalic surface ectoderm. Note where the position of the epibranchial placode is mapped in the young embryo. gl, sensory ganglion of the branchial nerve; ph, pharynx Fabric of the cranium Modern version of the germ-layer theory.
Оригинальная версия теории зародышевых слоёв была отклонена с открытием, что производная нервного гребня эктомезенхима вносит вклад в черепно-лицевой скелет. Считалось, что мезодермальная мезенхима является основным источником скелета позвоночных и то же самое значение придавалось головной мезодерме, т.к. она приравнивалась к сомитам туловища. Плакодное происхождение некоторых периферических ганглиев является др. причиной отказа от теории (rev. de Beer, 1958). Однако, мы всё ещё имеем тенденцию думать, что специфические клеточные клоны постоянно используются для специфического спектра клеточных типов или скелетных компонентов. В терминах современной биологии развития дух теории зародышевого слоя м.б. возрожден, из-за наших индуктивных пристрастий, что "морфологически гомологичные структуры (или стремятся быть) продуцируются из консервативных и ограниченных клеточных клонов".
Сегодня становится ясно, что обширная производная нервного гребня эктомезенхима в первую очередь оккупирует вентральные части позвонков головы, в противоположность более аксиально и дорсально локализованной головной мезодерме (Рис. 4, 9, 10; Noden, 1988). Эта вентральная эктомезенхима обнаруживается также у миног (Takio et al., 2004) и у эмбрионов hagfish (von Kupffer, 1900). Возникает несколько вопросов. Совпадают ли отдельные клеточные клоны (нервный гребень в сравнении с мезодермой) с анатомической конфигурацией краниума (висцерокраниум по сравнению нейрокраниумом)? Законсервирована ли конфигурация во время эволюции? В современной версии теория зародышевого слоя (клеточных клонов) вопрос д. звучать так: является ли морфологическая гомология черепа, постоянно возникающего из определенных специфических клеточных клонов, результатом специфических онтогенетических механизмов? Если нет, то имеются ли более подходящие морфологические подразделения черепа, которые бы соответствовали бы делениям клеточных клонов или типов клеток, таких как мезодерма и клетки нервного гребня? 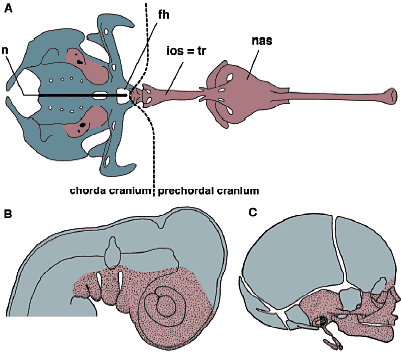 Fig. 10. Neural-crest-derivedts of the amniote cranium. (A) Origin of the 'neurocraniom' based on experiments on avian embryos reported by Couly et al., (1993). The mesodermal part of the cranium is colored blue, and the crest-derived part red. Note that the interface between the mesodermal and crest-derived parts of the neurocranium corresponds to the hypophysial foramen (fh) or the rostral limit of the notochord (n). (B) Distribution of the cephalic mesoderm (blue) and crest-derived ectomesenchime (red, stippled) in the chicken pharyngula by Noden (1988). Compare with Fig.9. The ectomesenchyme occupies the ventral half of the head, including the PAs and the premandibular region. (C) Results of avian chimeric experiments were extrapolated to the human perinatal skull based on Noden (1988). In his view, the cranial vault is thought to be of mesodermal origin. ios, interorbital septum; nas, nasal capsule; tr, trabecula Fig. 10. Neural-crest-derivedts of the amniote cranium. (A) Origin of the 'neurocraniom' based on experiments on avian embryos reported by Couly et al., (1993). The mesodermal part of the cranium is colored blue, and the crest-derived part red. Note that the interface between the mesodermal and crest-derived parts of the neurocranium corresponds to the hypophysial foramen (fh) or the rostral limit of the notochord (n). (B) Distribution of the cephalic mesoderm (blue) and crest-derived ectomesenchime (red, stippled) in the chicken pharyngula by Noden (1988). Compare with Fig.9. The ectomesenchyme occupies the ventral half of the head, including the PAs and the premandibular region. (C) Results of avian chimeric experiments were extrapolated to the human perinatal skull based on Noden (1988). In his view, the cranial vault is thought to be of mesodermal origin. ios, interorbital septum; nas, nasal capsule; tr, trabecula Neural crest versus head mesoderm Нейрокраниум локализуется в дорсальной части головы, энкапсулируя ЦНС, тогда как висцерокраниум поддерживает глотку со скелетным комплексом фарингеальных дуг (Рис. 3). Дермальная экзоскелетная и пред-сформированная хрящами эндоскелетная части ассоциируют с обоими компонентами (Рис. 3). Общепринято, что весь висцеральный скелет происходит из нервного гребня (Рис. 3, 4, 10). Следовательно, различие между нейрокраниумом и висцерокраниумом соответствует ли эмбриональным паттернам распределения и судьбам эктомезенхимы и мезодермальной мезенхимы?
Согласно Noden (1988) большинство краниофациальныха структур происходит из клеток нервного гребня, тогда как "нейрокраниум" является частично мезодермального происхождения. Из нейрокраниальный элементов полностью ethmoid (носовая капсула), часть клиновидной кости и часть отической капсулы возникают из клеток нервного гребня. Couly et al., (1993) более точно идентифицировал различия между производными нервного гребня и производными мезодермы частями основания черепа на уровне hypophysial отверстия (Рис. 10А). Эта граница соответствует месту, в котором трабекулярные хрящи прикрепляются к ростральному концу пара-хордального хряща или гребню, наз. 'crista sellaris' у некоторых амниот (de Beer, 1937; crista sellaris представляет собой задний край hypophysial отверстия и не является гомологом dorsum sellae у млекопитающих. Последнее является прямым производным орбитального хряща).
Распространенность мезодермального нейрокраниума соответствует ростральной границе хорды. Хорда и цефалическая мезодерма совместно заканчиваются рострально позади аденогипофиза. Поскольку мезодерма нуждается с сигналах, происходящих от хорды для хондрификации (Рис. 9-11) 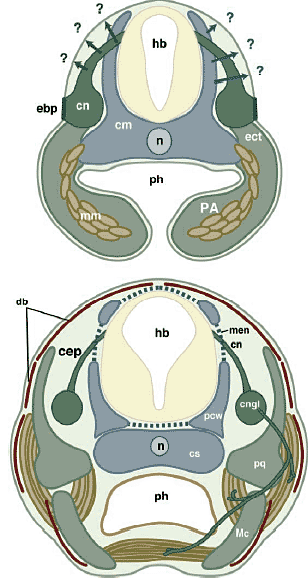 Fig.11. Cell lineages and topography. Anatomical and embryonic configuration of the vertebrate cranium at the level of the mandibular arch is duagrammatically illustated, based on Kuratani (2004a). The mesodermally derived neurocranium (blue) differentiates through induction by the nothchord (n) and the neuroectoderm (hindbrain; hb), and crest-derived cells form the cranial nerve (cn; dark green) and branchial skeletons (light green) in the pharyngeal arch (PA). In the completed cranium (below), the dermal bones (db; dark red) heve developed in superficial layers. Note that the dermdl bones in the skull roof can reasonably originate ether from crest cells or frpm the cephalic mesoderm (arrows). cep, cavum epiptericum; cm, cephalic mesoderm; cngl, cranial nerve ganglion; cs, centarl stem; ebp, epibranchial placode; ect, ectomesenchyme; Mc, Meckel's cartilage; men, meningeal membrane; mm, mandibular mesoderm; pcw, primary cranial wall derivwd from the cephalic mesoderm; pf, pharynx; pq, palatoquadrate. Fig.11. Cell lineages and topography. Anatomical and embryonic configuration of the vertebrate cranium at the level of the mandibular arch is duagrammatically illustated, based on Kuratani (2004a). The mesodermally derived neurocranium (blue) differentiates through induction by the nothchord (n) and the neuroectoderm (hindbrain; hb), and crest-derived cells form the cranial nerve (cn; dark green) and branchial skeletons (light green) in the pharyngeal arch (PA). In the completed cranium (below), the dermal bones (db; dark red) heve developed in superficial layers. Note that the dermdl bones in the skull roof can reasonably originate ether from crest cells or frpm the cephalic mesoderm (arrows). cep, cavum epiptericum; cm, cephalic mesoderm; cngl, cranial nerve ganglion; cs, centarl stem; ebp, epibranchial placode; ect, ectomesenchyme; Mc, Meckel's cartilage; men, meningeal membrane; mm, mandibular mesoderm; pcw, primary cranial wall derivwd from the cephalic mesoderm; pf, pharynx; pq, palatoquadrate.то Couly et al., (1993) назвали ростральную, происходящую из нервного гребня часть нейрокраниума 'prechordal cranium', который м. хондрифицироваться без индукции хордой. Очевидно, что такое эмбриональное разграничение черепа базируется на предположении, что определенные типы клеток постоянно нуждаются в одном и том же индуктивном механизме. Следовательно, боковые стороны черепа и основание м.б. подразделены на две части, соответствующие присутствию и отсутствию хорды, отражая различия в происхождении клеток (нервный гребень или мезодерма), а также различия в механизмах передачи сигналов, которые заставляют их дифференцироваться в скелетную ткань. Хотя происходящие из мезодермы скелетные элементы обнаруживаются в области, близкой к хорде, но Schneider (1999) установил, что если производную нервного гребня эктомезенхиму трансплантировать эктопически на место параксиальной мезодермы, предназначенной для образования орбитально-височной области, то она м. дифференцироваться в скелетные элементы, которые морфологически неотличимы от тех, что обычно продуцируются мезодермой. Т.о., клеточные клоны м. взаимозаменяться в определенном ограниченном онтогенетическом контексте независимо от классификации формируемых скелетных элементов. Более того, производная нервного гребня эктомезенхима скорее всего реагирет на сходные сигналы, которые способствуют скелетогенезу и облегчают собственно формирование паттерна скелетных элементов мезодермального происхождения. В нормальном развитии каждая часть отической капсулы, по-видимому, хондрифицируется посредством идентичного механизма индукции, т.к. капсула состоит из клеток нервного гребня и мезодермы. Следовательно, различия между передачей сигналов, специфичных для нервного гребня и специфичных для мезодермы, м.б. беспричинными (gratuitous) в узком смысле, а термины, такие как "нейрокраниум" и "висцерокраниум" м.б. в первую очередь ассоциированы с эмбриональными условиями (местами). Каждый тип клеток просто стремится занять специфическое положение в эмбрионе в результате, напр., специфических паттернов миграции клеток нервного гребня и оригинального распределения цефалической мезодермы, которые сильно ограничены во время филотипических стадий (эволюционно стабилизированы). Сходный феномен распознается для миогенных мезодермальных клеток. Хотя экспрессия некоторых миогенных генов регулируется клеточно-автономно, формирование морфологического паттерна клеток сильно зависит от эмбиологических условий (Borue, Noden, 2004). Этот феномен попирает современную версию теориии зародышевого листка. Согласно экспериментам Schneider генерация морфологического паттерна позвоночных в основном зависит от эпигенетических взаимодействий, которые базируются на топографически организованном морфологическом паттерне эмбриона, но не на программах целиком, ассоциированных с клеточными клонами. Origin of the dermal skull roof Проблема нервный гребень в сравнении с мезодермой касается и происхождения дермы свода черепа. Согласно морфологической концепции дермокраниум м.б. подразделен на висцеральный и нейральный компоненты и если задняя часть эндоскелетного нейрокраниума является мезодермального происхождения, то это верно и для дермальной компоненты этой части свода черепа (Рис. 3). Поэтому считалось, что эта часть дермы свода черепа является мезодермального происхождения. Однако, Couly et al., (1993) показали. что эти скелетные элементы также происходят из нервного гребня. Хотя большая часть эктомезенхимы оккупирует вентральную часть эмбриональной головы,, места, на которых дифференцируются дермальные элементы соответствуют дорсо-латеральным путям миграции, характерным для клеток цефалического нервного гребня (Рис. 11). Соответственно, или клетки, происходящие из нервного гребня или цефалической мезодермы, д. дифференцироваться в эти скелетные элементы.
Анализ трансгенных мышей показал, что имеется передне-заднее различие в дермальных костях между теми, что произошли из клеток, которые однажды были активированы промотором Wnt1 (возможный клональный маркер для клеток нервного гребня) и теми, что произошли из клеток , которые не были активированы IJiang et al., 2002; Morris-Kay, 2001). Эта проблема, которая касается наиболее поверхностно локализованных скелетных элементов, всё ещё не решена. Возможно, что гомологи дермальных элементов развиваются из разных клеточных клонов в каждой группе животных. Здесь снова топографически д.б. только один фактор, навязывающий онтогенетические ограничения, продуцируя сигналы для морфологических гомологов. В этом контексте гомологи дермальных костей были описаны у водных видов для морфологии системы латеральных линий (Jarvik, 1980; Starck, 1980). Подобно формированию отической капсулы, формирование паттерна дермальных костей также возможно м.б. эпигенетическим событием, не связанным жестко с любым специфическим клеточным клоном. Evolution and development of the viscerocranium Cephalic Hox code and branchiomerism
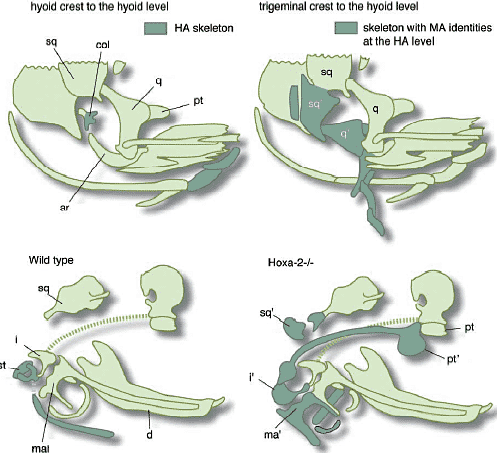
Если скелеты РА действительно являются серийными гомологами, то как они м. дифференцироваться в специфические морфологии в соответствии со своей позицией? Noden (1983) показал, что когда нервный гребень, предназначенный занять мандибулярную дугу (РА1), трансплантировался на уровень гиоидной дуги (РА2)(приблизительно на уровне 4 ромбомера) хозяина, то некоторые скелетные элементы в РА2 развивались с мандибулярными качественными особенностями, такие как qiadrate и articular, скорее, чем как гиоидные скелетные элементы, такие как colimnella auris и retroarticular process, которые обычно образуются в этой дуге (Рис. 12А).
  Нох код также функционирует и в РА эктомезенхиме. У всех изученных gnathostome эмбрионов PG2 Hox гены экспрессируются в РА2 и каудальнее её, PG3 гены в РА3 и каудальнее и т.д. (Рис. 4ж Hunt et al., 1991фби)Нет Нох генов, экспрессируемых в РА1 и дифференцировка челюстей, по-видимому, специфицируется в отсутствие Нох транскриптов в эктомезенхиме ('Hox-code default state') в этой дуге. Показано, что agnathans м. обладать сходным тем же Нох кодом, состоящим из PG2 и PG3 от РА1 через всю РА3 эмбриональной глотки (Takio et al., 2004; Kuratani, 2004b; Cohn, 2002).
Онтогенетическая функция 'cephalic Hox code' заключается в спецификации, по крайней мере, морфологии РА2, как оппозитной РА1. Нарушение экспрессии Ноха-2, обычно обнаруживаемой с РА2 и каудальнее, ведет к трансформации РА2, которая начинает частично походить на РА1 (Рис. 12). Напротив, избыточная экспрессия Ноха-2 вызывает трансформацию РА1 в направлении РА2 (Pasqualetti et al., 2000; Grammatopoulos et al., 2000). В этих экспериментах, следовательно, РА1 м РА2, по-видимому. представляют собой эквивалентные онтогенетические единицы, которые м. изменять свои онтогенетические судьбы, когда разные позиционные значения экспериментально накладываются на них, создавая онтогенетическую основу для бранхиомерной трансформации.
Схема цефалического Нох кода частично согласуется с классической концепцией трансформации бранхиальных дуг. Родоначальные позвоночные были использованы, чтобы получить серии недифференцированных РА и каждая РА постепенно приобрела свою специфическую программу дифференцировки в ходе эволюции в зависимости от свойи позиционных значений. В отличие от ситуации, наблюдаемой в эволюции формулы позвоночных, регуляция Нох кода в РАs, по-видимому, не менялась в ходе эволюции. Следовательно, м. обозначать эквивалентные дуги у разных животных одними и теми же именами, напр., "мандибулярная", "гиоидная". Важно, что родоначальник с недифференцированной серией РА, по-видимому, чисто теоретически обладает качественными особенностями мандибулярной, гиоидной, т.е. качественные особенности бранхиальных дуг, по-видимому, уже присутствуют у общего предшественника миног и gnathostomes (Takio et al.,2004; Kuratani et al., 2004b). Более того, происхождение челюстей, по-видимому, происходит в результате сложного сдвига в тканевых взаимодействиях, а не в результате простой трансформации мандибулярной дуги ( Kuratani et al., 2001; Shigetani et al., 2002; rev. Kuratani et al., 2004a; Lee et al., 2004; Verny et al., 2004). 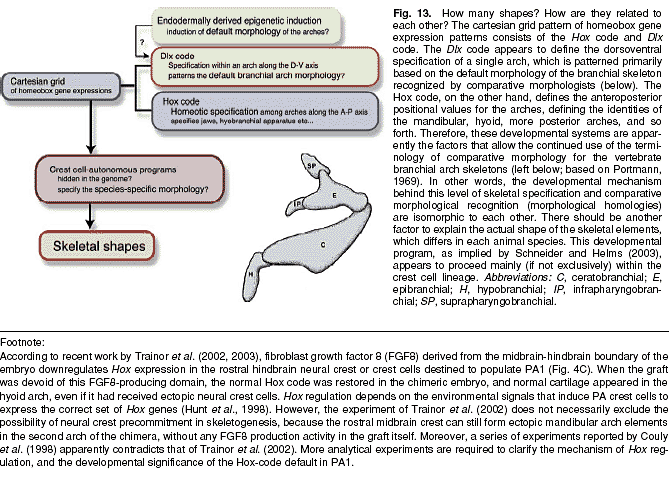 Во всяком случае Нох код является не более чем онтогенетической системой, с помощью которой задаются позиционные значения для каждой дуги, а 'Hox-code default' не обязательно средний (mean) 'прототип PAs' во всех смыслах (Рис. 13). Мандибулярная дуга у позвоночных обычно наиболее сильно разнообразна из всех дуг. В сравнительной морфологии прототипическая морфология РА идентифицируется в форме пост-отических бранхиальной дуги, в которой имеется определенное количество хрящевых элементов, обычно идентифицируемых у многих gnathostomes и даже форма скелета мандибулярной дуги м. рассматриваться как модифицированная версия этого паттерна (Portman, 1969; Jarvik, 1980; Kuratani, 2004b). Но этот паттерн не представляет 'ancestral shape' скелета дуги, т.к. подобного скелетного паттерна не найдено у agnathans. Jaw and trabecula Онтогенетическое значение 'Hox-code default state' также остается неясным. Как показано, с помощью разрушения Ноха-2 удвоенные морфологические качественные особенности наблюдаются только в проксимальной части мандибулярной дуги и Hox-code default state не формирует, по-видимому, паттерна более ростральной части скелета мандибулярной дуги. Др. гомеобоксный ген Otx-2 м.б. ответственен за формирование паттерна этой области, т.к. гаплонедостаточность по этому гену вызывает спектр редукций в проксимальной части РА1 (Matsuo et al., 1995). Др. словами, и экспрессия Otx-2 и Нох-код default состояние вместе формируют паттерн РА1 комплементарно (кум. Kuratani et al., 1997). Интересно, что паттерны экспрессии у миноги Otx-2 и некоторых Нох генов выглядит очень сходным с таковым у gnathostomes (Takio et al., 2004), указывая тем самым, что молекулярное кодирование РА1 устанавливается очень рано в истории позвоночных, в зависимости от того, развиваются или нет челюсти у животных. Являются ли эти два домена настоящими компартментами и где в мандибулярной дуге проходит граница, являются нерешенными вопросами.
Довольно проблематичным является присутствие более рострально локализованной эктомезенхимы, также происходящей из trigeminal клеток нервного гребня, которая не экспрессирует каких-либо Нох генов, ростральнее мандибулярной дуги. Это соответствует области, где дифференцируется 'prechordal cranium' как предполагает Couly et al., (1993)(Рис. 9; Kuratani et al., 1997; 2004). Т.о., прехордальный краниум м. рассматриваться не только как производная нервного гребня ростральная половина нейрокраниума, но и как скелетная часть, дифференцирующаяся из ростральной половины клеток trigeminal нервного гребня, которые д. называться клетками 'premandibular' нервного гребня (Рис. 6).
В некоторых классических концепциях термин "премандибулярный" используется, чтобы отразить присутствие др. фарингеальной дуги впереди мандибулярной дуги у родоначальников позвоночных, что принято не всеми. Палочковидной формы скелет, трабекулярный хрящ, напоминают фарингеальную дугу. Более задние мезодермальные нейрокраниальные элементы, парахордальные хрящи также парные и палочковидные (о trabecula cranii см. de Beer, 1931, 1937; о хряще того же названия у миног см. Johnes, 1948; Kuratani et al., 2001; 2004). Идея "премандибулярной дуги" целиком идеалистична (о реинпретации agnathan fossil доказательствах см. Janvier, 1996; Kuratani, 2005). Он оценивается как гипотетический головной сегмент, к которому принадлежат ophthalmic нерв и премандибулярная мезодерма, как противовес довольно четкому мандибулярному сегменту, который включает мандибулярную дугу, максилло-мандибуляоную часть тройничного нерва и мандибулярную головную полость у акул. Очевидно, что эта схематическая интерпретация головы позвоночных базируется на предположении, что гипотетические мезодермальные сегменты (опровергнутые выше) и branchiomtric фарингеальные дуги ассоциированы др. с др. один-к-одному (Kuratani, 2003). Если мы примем такую унифицированную сегментную схему для головы позвоночных в контексте последней эволюционной онтогенетической теории, то д.б. одно общее генеративное ограничение, которое затрагивает сегментную организацию фарингеальных дуг, мезодермы и возможно ромбомеров вместе, в качестве особого вышестоящего онтогенетического события. Однако, не идентифицировано такого онтогенетического события. 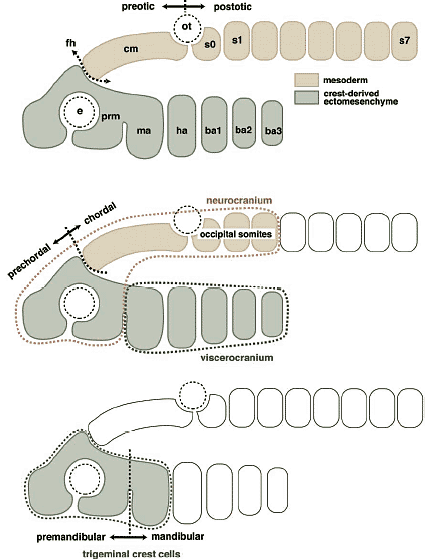  Игнорирование сегментной организации головы позвоночных без сомнения касается большой эктомезенхимной части клеток trigeminal нервного гребня , ростральнее РА1 клеток нервного гребня (Рис. 14). В раннем эмбриогенезе кур клетки trigeminal нервного гребня формируют непрерывный слой клеток без границ. Однако, благодаря индуктивному действию FGF8, локализованного в вентральной части эктодермы, каудальня половина клеток специфицируется как клетки мандибулярного нервного гребня, остальные определяются как клетки премандибулярного нервного гребня (Shigatani et al., 2000; rev. Kuratani, 2005). Исходя из регионализованного распределения клеток нервного гребня (Kontges, Lumsden , 1996) клетки премандибулярного нервного гребня, по-видимому, происходят из нервного гребня между передним мозгом и ростральной частью среднего мозга (Osumi-Yamashita et al., 1994). Однако, это не означает, что эти клетки пре-детерминированы становиться премандибулярными структурами в премиграторном состоянии. Следовательно, предполагаемые премандибулярные скелетные структуры, происходящие из нервного гребня, такие как trabecula и носовая капсула, дифференцируются после премандибулярной-мандибулярной спецификации клеток trigeminal нервного гребня. Более того, подобно остальной части РА скелета, премандибулярные скелетные элементы нуждаются в присутствии энтодермы, но не хорды для хондрификации.
Если они лишены некоторых онтогенетических и морфологических признаков. необходимых им для их т.наз "фарингеальных дуг", то премандибулярная и РА эктомезенхима сходны по способу своей хондрификации. Напр., премандибулярная часть краниума зависит от взаимодействия с энтодермой (Couly et al., 2002), но не с передним мозгом и эктодермальным эпителием (Noden, 1978). Как отмечалось выше, эти скелетные элементы называются "прехондральным" краниумом в противовес "хордальному" краниуму в терминах отдельных клеточных клонов и механизмов спецификации передачи сигналов скелетогенеза (Рис. 10, 13). Рассматривая ещё более ранний эмбриогенез, когда непрерывная эктомезенхима вторично регионализуется посредством тканевых взаимодействий в сегментированную глоточную область и частично ростральнее её (вне сегментного контекста), то мы м. распознать различия между "премандибулярной" (вообще-то более правильно назвать "пре-фарингеальной") и "мандибулярной" эктомезенхимой (Рис. 14) Хотя первая интегрируется в нейрокраниум в функциональном смысле и одновременно представляет собой наиболее ростральный компонент краниума, производного нервного гребня, который зависит от присутствия цефалической энтодермы (см. ниже). Бранхиомернаная фарингеальная эктомезенхима дуг в этом отношении д. скорее всего рассматриваться как вторично сегментированная часть головы позвоночных, которая заполняется клетками цефалического нервного гребня (Рис. 1 из Kuratani, 2005). Присутствие эктомезенхимы, производной нервного гребня, per se не обязательно д. приводить к присутствию бранхиомерных сегментов, поэтому не обязательно предполагать, что у предшественников позвоночных цефалическая эктомезенхима была полностью сегментирована. Cartesian grid of homeobox gene expression and enviromental cues Подобно Нох коду, который функционирует вдоль передне-задней оси, гены Dlx экспрессируются сходным гнездовым паттерном у мышей и возможно у др. gnathostome эмбрионов. У мышей Dlx1 и Dlx2 экспрессируются скорее повсеместно в РА эктомезенхиме, тогда как экспрессия Dlx5 и Dlx6 ограничена вентральной половиной PAs (Рис. 4). Более того, Dlx3 и Dlx7 экспрессируются только в дистальных (вентральных) кончиках PAs, завершая гнездовой паттерн экспрессии генов Dlx, обозначаемый как Dlx code (Depew et al., 2002). Эти гены. по-видимому, формируют паттерн скелета дуг вдоль дорсо-вентральной оси, т.к. одновременное нарушение генов Dlx5 и Dlx6 дает зеркальные удвоения верхнечелюстных элементов вместо нижнечелюстных элементов (Beverdam et al., 2002; Ozeki et al., 2004). Следовательно, в терминах онтогенетического программирования, верхнечелюстная морфология являет собой default качественные особенности, а формирование паттерна нижних частей челюстей м.б. связано со вторичным привлечением вентрально экспрессирующихся транскрипционных факторов (Depew et al., 2002). Возможно затем, что такой Dlx код является обязательным условием для дорсо-вентральной спецификации РА скелета, включая челюстных (rev. Schilling, 2003), а у миног с их дорсовентральным симметричным скелетом фарингеальных дуг, он еще не достигнут на этой стадии эволюции (Тушвуке уе al., 2001; Myojin et al., 2001; rev. Schilling, 2003; Dhigetani et al., 2005).
Интересно, что у Dlx5/Dlx6 с двойным нокаутом и у Ноха-2-мутантных мышей удвоенные скелетные элементы обнаруживают симметричные паттерны в отношении исходных скелетных элементов. Хрящи basket фарингеальных дуг у миног обнаруживают дорсо-вентральную симметрию и мы выявили, что фарингеальные карманы находтся на оси этой симметрии. Это совпадение указывает на индуктивную функцию фарингеальной энтодермы на формирование паттерна скелета, это указывает на то, что Нох и Dlx коды являются простыми системами, которые обеспечивают позиционные сигналы, а не формируют очертания скелета. Это м. б. также объяснить, почему Нох-негативный нервный гребень всегда продуцирует одну и ту же часть скелета мандибулярной дуги (артикуляция челюсти), если помещается на уровень предназначенный для конца второй дуги (Couly et al., 1998).
Couly et al. (2002) удаляли каждую часть ростральной энтодермы со стадии 8 у эмбрионов кур и обнаружили, что разные части производного нервного гребня краниального скелета теряются в каждом случае в зависимости от передне-заднего уровня, с которого была удалена энтодерма. Наиболее ростральная энтодерма или preoral кишка была необходима для хондрификации прехордального (премандибулярного) краниума, а несколько более каудальный уровень энтодермы для ростральных кончиков скелета мандибулярной дуги и т.д. Сходным образом, эктопически имплантированная энтодерма индуцирует скелетные элементы со специфическими качественными особенностями и ориентацией, в зависимости от происхождения и ориентации трансплантированной энтодермы. Эти эксперименты указывают на то, что схематическое представление о предопределении нервным гребнем качественных особенностей скелета м.б. передвинуто на энтодермальный слой, которой организован как решетка (lattice)? предопределяемая передне-задней и дорсо-вентральной осями. Однако, всё это не так просто из-за того, что (1) инактивация функции Нох генов во второй дуге все еще приводит к трансформации скелета гиоидной дуги, придавая им качественные особенности мандибулярной дуги, где еще происходят обычные энтодермой предопределяемые индуктивные события: и (2) как показано Wagner (1959) видо-специфичная форма нервным гребнем предопределяемого скелета, по-видимому. кодируется в нервном гребне ещё до миграции, а не условиями хозяина. включая и энтодерму.
В ответ на первый вопрос мы м. предсказать, что энтодермально определяемые сигналы м.б. действительно теми же самыми для эктомезенхимы мандибулярной дуги и гиоидной дуги ( но см. Ruhin et al., 2003). Функция Нох д. "модулировать" ниже стоящий процесс дифференцировки, в результате чего формируются скелеты с отличающимися качественными особенностями. В отношении второй проблемы, по-видимому, заставляющей нас разделить концепцию "формы" или "identity" скелета из нескольких отличающихся уровней или типов. Энтодерма посылает в направлении клеток нервного гребня сигналы. которые предопределяют морфологические качественные особенности, такие как 'quadrate' или 'articular', которые обычно обнаруживаются у разных групп животных. Клетки нервного гребня транслируют эти сигналы, используя свои собственные геномы для предопределения действительной формы, которая уникальна для каждой группы животных. Если содержание энтодермально испускаемых сигналов до некоторой степени сходно со структурой (framework) на уровне сравнительной морфологии, реакция клеток нервного гребня д.б. более сходной с действительной анимальной формой, которая д.б. более или менее клеточно автономной в клонах нервного гребня. Присутствие этих разных уровней морфогенеза согласуется с уникальным экспериментом, осуществленным Schneider и Helms (2003). Whot determines the shape? How many types of shapes? Цефалический Нох код в фарингеальных дугах впервые появляется, чтобы сравнивать ранние данные экспериментально эмбриологии. Считается, что скелетная форма предетерминируется премиграторными клетками нервного гребня. В контексте "скелетных качественных особенностей" это предетерминирование нервного гребня обычно через чур упрощается, а позиционные значения и видо-специфичная морфология часто не согласуются. Напр., межвидовые трансплантации нервного гребня между Tritrus и Bombina lf.n скелет с донорской морфологией у химер (Wagner, 1959). "tcgthbvtyns Noden (1983), с др. стороны, хотя и не имеют непосредственного отношения к этому вопросу, а к позиционным значениям эктомезенхимы фарингеальных дуг вдоль передене-задней оси.
Schneider и Helms (2003) обменивали премиграторные цефалические нервные гребни между эмбрионами уток и перепелов, птиц с разной черепно-лицевой морфологией (Рис. 15).
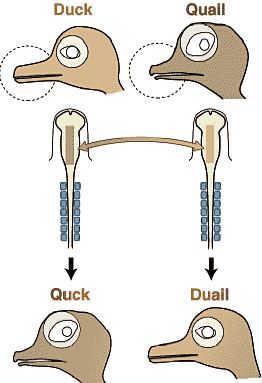 Fig. 15.Does species-specific morphology reside in the crest cells? Based on the experiment by schneider and Helms (2003). When the cephalic neural crest is exchanged between the duck and quail, the morphology of the ectomesenchymal part of the head develops into the identity of the crest donor species. Because the developmental patterning of the crest-derived cells ewquires the embryonic enviroment including the endoderm and ectoderm (host tissue), this experiment implies there is a developmental process that generates species-specific traits associated with the crest cell lineage, more or less independet of the epigenetic, tissue interactions. Fig. 15.Does species-specific morphology reside in the crest cells? Based on the experiment by schneider and Helms (2003). When the cephalic neural crest is exchanged between the duck and quail, the morphology of the ectomesenchymal part of the head develops into the identity of the crest donor species. Because the developmental patterning of the crest-derived cells ewquires the embryonic enviroment including the endoderm and ectoderm (host tissue), this experiment implies there is a developmental process that generates species-specific traits associated with the crest cell lineage, more or less independet of the epigenetic, tissue interactions. Интересно, что форма, генерируемая у химер всегда была более сходна с той, что у вида донора, чем хозяина. Следовательно, как и в экспериментах Wagner (1959) "форма" химерного скелета базируется в клетках нервного гребня. Эмбриональные условия тканей хозяина, по-видимому. посылают те же самые индуктивные сигналы, но клетки нервного гребня, которые получают такие сигналы м. только отвечать на базе собственного генома в своих ядрах. Клетки нервного гребня перепела не знают, как собираться в 'duck quadrate',когда они получают приказ от эмбриональных условий утки "делать quadrate". Эксперименты Schneider и Helms (2003), как и Wagner (1959) указывают на то, что м.б., по крайней мере, два способа делать форму хряща: "видо-специфическая видимая форма" и "эквивалентные качественные особенности" скелетных элементов, т.е. два разных скелетных элемента у двух разных видов животных с одним названием в сравнительной морфологии (Рис. 13). Следует помнить, что концепция морфологической гомологии не нуждается в какой-либо сходстве действительной формы или функции, но д. базироваться на эквивалентном относительном положении в общем плане тела. И снова "общее топографическое положение" означает идентичную эпигенетическую индукции обеих тканей. Если такое взаимодействие является эволюционно фиксированным и неизменным, то эта неизменность д. распознаваться как онтогенетическое ограничение, которое генерирует "морфологическую гомологию". Это близко к идее, что филотипическая стадия развития животных имеет тенденцию быть законсервированной посредством сложной сети глобальных взаимодействий и что эмбриональные паттерны, обнаруживаемые на филотипических стадиях являются источником наиболее глобальной гомологии, которая предопределяет план тела животных phyla.
Важно, что эти разные уровни концепции м.б. классифицированы с помощью соотв. образом организованных экспериментов и точного понимания механизмов формирования онтогенетических паттернов.
Conclusions and perspectives The experiment of Noden (1983) involving transplantation of the mandibular crest to the level of the hyoid, and that of Trainor etal. (2002), which implies an epigenetic function of midbrain-hindbrain boundary must be reconciled with that of Couly et at. (1998) in the context of regulation of the cephalic Hox code and its maintenance (or restoration). Undoubtedly, there is a certain level of environmentally derived signals that maintains or upregulates the Hox gene expression, as predicted by Hunt et al. (1998), who rotated the whole hindbrain along the anteroposterior axis and had restored the correct Hox code. Simultaneously, when grafted crest cells formed a large cell population, there would have been a community effect that would maintain the same original Hox gene expression under a varied environment, leading us to believe that Hox regulation in the crest is, at least as a phenomenon, precommitted at 'the premigra-tory state along the neuraxis.
Importantly, the segmental deployment of crest cells and the expression of Dlx and Hox genes are spafiotempo-rally highly organized at the stage of phylotype, on which both the developmental specification and evolutionary changes are dependent No doubt the acquisition together of such an organized embryonic pattern and gene expression patterns is one of the most crucial factors in the morphogenetic events of the vertebrate cranium. It is highly conceivable that such patterns were necessarily stabilized through evolutionary selection; &e developmental mechanism and genes could change without altering the patterns generated. Furthermore, the pseudosagmental patterns in the vertebrate phylotypic cranium may be the most important developmental factor (developmental constraint) in the morphological homology of skeletal elements. This pattern is obtained secondarily in embryonic development, and is not present in very early embryos. In this context, it is worth mentioning that the results of mapping studies performed in the cephalic mesoderm of two different stages of chick embryos by Couly et al. (1992) and Noden (1988) differ greatly (Fig. 6). The fate map at the late neurula is reminiscent of Goodrich's segmental theory, whereas such a pattern is not yet established when the fate mapping is performed at earlier stages.
In conclusion, comparative embryology of the vertebrate cranium has shown the presence of a developmentally constrained pattern of embryos, and the resulting tissue interactions that give rise to certain specific patterns of skeletal elements. We can now identify the types of interactions and cell movements that are crucial in the generation of certain specific morphological patterns, and the developmental and evolutionary contexts that must be addressed to better understand craniogenesis. With the molecular developmental and genetic techniques available to us, the longstanding question of the Vertebrate head' has now reached its final stage of resolution.
|